
Traduit en Français par François Mériaux (fm@osteo-animalier.com)
La grande majorité des publications scientifiques consacrées à l’ostéopathie est rédigée en anglais et demeure, de fait, peu accessible à de nombreux praticiens francophones. L’ objectif de cette traduction est donc de leur rendre ces données plus facilement disponibles.
Basée sur l’analyse de plus de 150 publications scientifiques, cette revue propose un état des lieux des connaissances concernant le MRP. Elle revisite les fondements historiques du modèle développé par Sutherland, examine les données expérimentales disponibles et explore plusieurs hypothèses physiologiques susceptibles d’expliquer les phénomènes observés en pratique.
Ainsi, cette traduction s’adresse aussi bien aux étudiants qu’aux praticiens souhaitant approfondir leur compréhension de l’ostéopathie fluidique et des débats scientifiques qui l’entourent.
J’espère qu’elle contribuera à nourrir la réflexion, encourager l’esprit critique et favoriser des échanges constructifs autour de l’évolution des modèles explicatifs de l’ostéopathie crânienne.
https://doi.org/10.32388/S6IGL0
Résumé
L’Impulsion Rythmique Crânienne (IRC), traditionnellement attribuée au Mécanisme Respiratoire Primaire (MRP), désigne un rythme perçu par certains ostéopathes lors de la palpation et utilisé en pratique clinique à des fins diagnostiques et thérapeutiques. Malgré son utilisation répandue, les bases physiologiques de ce phénomène demeurent incertaines.
Le modèle théorique proposé par William Garner Sutherland en 1939 a profondément influencé la compréhension du MRP/IRC. Cependant, les progrès réalisés en physiologie, en neurosciences et en biomécanique ont remis en question plusieurs de ses postulats fondamentaux. Depuis les années 1990, différentes hypothèses alternatives ont émergé, notamment celles mises en évidence par plusieurs revues systématiques récentes.
Dans cette revue narrative, nous réexaminons d’abord les cinq piliers historiques du modèle de Sutherland à la lumière des connaissances scientifiques contemporaines. Après avoir mis en évidence les limites de ce cadre théorique, nous explorons différents mécanismes physiologiques susceptibles de contribuer au phénomène MRP/IRC, notamment la vasomotricité, l’activité du système nerveux autonome, la variabilité de la fréquence cardiaque, la microcirculation, les dynamiques lymphatiques ainsi que le comportement de la matrice extracellulaire.
L’hypothèse la plus plausible est que le MRP/IRC constitue un phénomène émergent résultant de l’interaction et de l’entrainement partiel de multiples oscillations physiologiques, en particulier celles dépendantes de la régulation autonome, telles que les ondes de Traube-Hering-Mayer, les rythmes respiratoires et les variations cardiovasculaires. Ces interactions pourraient induire des modifications rythmiques des propriétés viscoélastiques de la matrice extracellulaire et du réseau fascial.
Enfin, nous discutons de l’intérêt clinique du MRP en pratique ostéopathique. Bien que le MRP/IRC puisse constituer un retour palpatoire utile pour le praticien, son interprétation doit rester prudente en raison de l’influence de nombreux facteurs physiologiques et perceptifs.
Cette revue souligne la nécessité d’un changement de paradigme dans la compréhension du MRP/IRC. Elle plaide en faveur d’un modèle fondé sur les connaissances scientifiques actuelles et soutient l’adoption d’une terminologie reflétant plus fidèlement l’état contemporain des connaissances.
Introduction
Le terme Mécanisme Respiratoire Primaire (MRP) est couramment utilisé par certains ostéopathes pour décrire la sensation palpatoire d’un rythme physiologique à la surface de la peau [1].
Il peut être utilisé pour leur diagnostic et leur traitement.
Il est décrit comme ayant deux phases, une phase d’expansion (inspiration) et une phase de contraction (expiration) [2][3].
Il est différent de la respiration puisqu’il reste perceptible lorsque le patient retient sa respiration [4].
Historiquement, l’ostéopathe qui l’a décrit en premier, William Garner Sutherland (1873-1954), a eu l’intuition que les sutures crâniennes étaient faites pour le mouvement et a imaginé un modèle basé sur cinq principes : la mobilité inhérente du système nerveux central et de la moelle épinière ; la fluctuation du liquide céphalo-rachidien (LCR) ; les membranes de tension réciproque ; la mobilité articulaire des os du crâne ; le mouvement involontaire du sacrum entre les os iliaques [2][3].
Sutherland a choisi le terme « Mécanisme Respiratoire Primaire » (MRP) pour mettre en évidence plusieurs aspects clés de son concept.
Le mot mécanisme indique que le MRP est un phénomène physiologique organisé, où plusieurs éléments interagissent de manière coordonnée.
Le terme respiratoire a été choisi par analogie avec la respiration pulmonaire, mais il se réfère à un processus différent.
Pour Sutherland, le MRP représentait une pulsation rythmique autonome dans le corps, essentielle à la vie et présente même en l’absence de respiration pulmonaire [2].
Il voulait souligner que le corps possède un mouvement périodique interne, indépendant des poumons, animant les tissus et les fluides.
Le mot primaire signifie que ce mécanisme serait présent dès le début de la vie embryonnaire et constituerait une fonction vitale fondamentale.
De nombreux ostéopathes utilisent aujourd’hui le terme MRP pour décrire les micromouvements avec lesquels ils travaillent. À noter qu’en France, un glissement sémantique est observé : de nombreux ostéopathes utilisent le terme « mouvement » plutôt que « mécanisme » lorsqu’ils font référence au MRP.
Cependant, pour être précis et rester fidèle à son sens d’origine, le terme Impulsion Rythmique Crânienne (IRC) devrait être utilisé lorsqu’on fait spécifiquement référence à la sensation palpatoire de ce rythme à la surface de la peau.
Cette terminologie a été introduite en 1961 par Woods et Woods, qui ont perçu un rythme à la surface de la tête.
Ils l’ont relié au MRP de Sutherland et ont tenté, sept ans après sa mort, de définir une plage de fréquence normale, ce que Sutherland lui-même n’avait jamais fait [5].
Pour certains ostéopathes, l’IRC et le MRP représentent le même mouvement, ressenti respectivement au niveau du cuir chevelu et dans le reste du corps.
La confusion entre ces deux termes est fréquente [6], sans être problématique dans la mesure où l’IRC est généralement considérée comme une manifestation du MRP, cette assimilation relevant ainsi essentiellement d’une simplification.
Le MRP reste l’explication historique de ce rythme particulier, qui est l’objet de cette revue [7][8].
Les cinq phénomènes sous-jacents au MRP ont été décrits pour la première fois par Sutherland dans The Cranial Bowl (1939) et ont ensuite été largement diffusés par son élève Harold Magoun, qui a publié Osteopathy in the Cranial Field en 1951.
Magoun a ensuite cherché à structurer les enseignements de Sutherland, en les rendant plus accessibles et reproductibles par la formalisation de techniques et de protocoles [2][3].
Cependant, Sutherland était quelque peu réticent à cette formalisation biomécanique, craignant qu’elle ne limite une compréhension plus profonde de son approche.
Alors qu’il considérait le mouvement crânien comme un processus holistique influencé par de multiples facteurs, Magoun, dans son effort de popularisation de ces idées, risquait de réduire cette complexité à un ensemble prédéfini de pratiques, en décrivant des mouvements et des techniques spécifiques.
Cette divergence a créé certaines tensions, Sutherland craignant que l’essence de son travail ne soit perdue [2][3].
Néanmoins, tel est l’héritage dont nous disposons aujourd’hui : les ostéopathes perçoivent un rythme supposé être associé aux mouvements des os du crâne et à d’autres phénomènes, et c’est précisément cette hypothèse que nous cherchons à réévaluer.
Avec ces clarifications à l’esprit, nous avons choisi d’associer systématiquement les termes MRP et IRC afin de nous aligner sur l’usage courant, en faisant référence au rythme typique perçu par les ostéopathes.
Le MRP/IRC est défini par ses trois composantes : amplitude, rythme et force.
Si ces trois composantes sont correctes, l’ostéopathe peut conclure que le patient, ou au moins la zone sur laquelle il se concentre, est bien équilibré.
Certains ostéopathes pensent que le MRP/IRC peut être utilisé comme un outil pour vérifier l’efficacité de chaque traitement ostéopathique, en le contrôlant après chaque technique appliquée jusqu’à ce qu’il soit suffisamment bon pour marquer la fin de la séance.
Le MRP/IRC refléterait alors l’harmonie et la vitalité du corps.
L’amplitude devrait être large et symétrique.
La force est considérée comme correcte lorsqu’elle est perçue comme puissante.
Bien qu’en général le rythme soit considéré comme normal entre 6 et 12 cycles par minute (cpm), la littérature rapporte de grandes variations allant de 3 à 22 cpm [9].
La Figure 1 montre l’étendue des fréquences décrites dans la littérature, qui ne sont pas facilement attribuables à un rythme physiologique identifié.
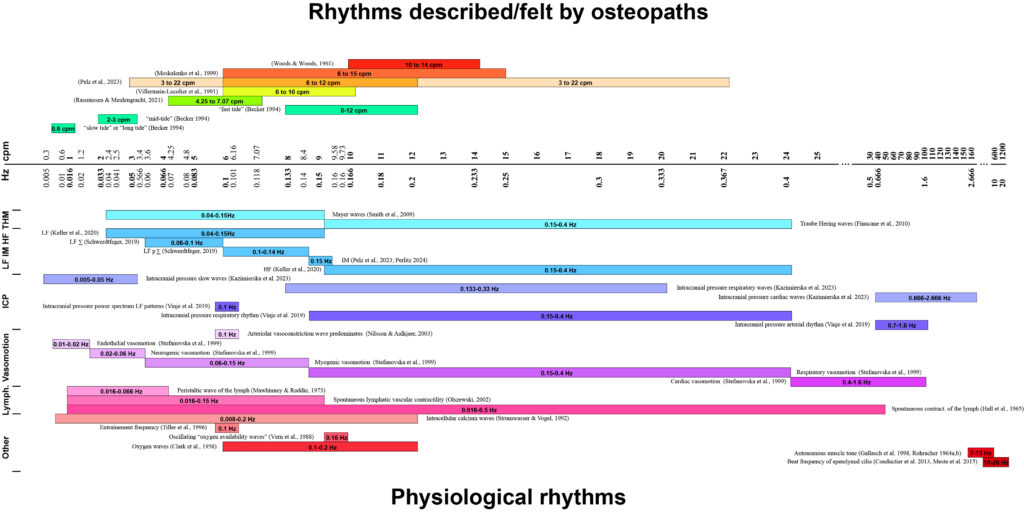
Figure 1. La présentation suivante exposera les différents rythmes et fréquences (exprimés en Hz et en cpm) décrits par les ostéopathes, en comparaison avec ceux des rythmes physiologiques. Ces rythmes seront regroupés dans les catégories suivantes : ondes de Traube Hering et Mayer (THM) ; basse fréquence (LF) ; fréquence intermédiaire (IM) ; haute fréquence (HF) ; onde de pression intracrânienne (ICP) ; vasomotion ; contractilité lymphatique spontanée (Lymph.) et autres. Pour Pelz et al. [8], l’intervalle de fréquence compris entre 3 et 22 Hz représente l’intervalle total des mesures, les intervalles plus restreints entre 6 et 12 Hz représentant la plage des valeurs les plus fréquemment retrouvées.
Ainsi, les différents rythmes perçus pourraient dépendre du couple praticien/patient et varier au cours du temps [10][11][12][2][13][14][15][5], rendant alors la mesure objective et la comparaison difficile, ce qui pourrait expliquer la faible reproductibilité intra- et inter-examinateur observée dans la littérature.
Par ailleurs, différentes études peuvent décrire des rythmes distincts sous une même terminologie.
Fait intéressant, des rythmes appartenant à la gamme de fréquence du MRP ont également été mis en évidence par des instruments de mesure, suggérant une possible objectivation du phénomène [10].
Dans une étude récente, Rasmussen et Meulengracht [4] ont démontré l’existence d’un troisième rythme physique (différent des rythmes cardiaque et respiratoire) détecté au niveau de la tête humaine par leur appareil, avec une moyenne de 6,16 cycles/minute (4,25–7,07).
Cependant, l’outil de mesure, bien que validé sur le plan métrologique dans leur laboratoire (Annexe A. Données supplémentaires dans [4]), présente un biais important, similaire à celui de l’outil utilisé par Frymann en 1971 : les mouvements de la tête générés par la ventilation ne sont pas contrôlables [16].
Nelson et al. [17][18][19][20], et plus récemment Pelz et al. [9], ont montré une corrélation statistiquement significative entre les mesures du flux sanguin cutané et la palpation de l’IRC, montrant que les manipulations crâniennes affectent ces paramètres objectifs.
Considérant l’existence d’études sérieuses suggérant que le MRP/IRC pourrait exister et qu’un praticien entraîné pourrait le percevoir, nous avons récemment réalisé une revue systématique structurée de la littérature dans MEDLINE, Science Direct et la Cochrane Library sur ce sujet afin de décrypter les différentes hypothèses des mécanismes physiologiques supposés sous-tendre le rythme MRP/IRC [1].
Cette revue systématique et la Figure 1 montrent que la question de la base physiologique de ce rythme reste controversée.
Le modèle initial proposé par Sutherland n’est plus compatible avec les connaissances physiologiques actuelles, et les mécanismes sous-jacents à ce phénomène restent incertains.
Les résultats de notre précédente revue systématique ont mis en évidence la nécessité d’un changement de paradigme et d’une évaluation et communication plus rigoureuses d’un modèle en accord avec l’évolution des données scientifiques.
C’est dans cette perspective que s’inscrit la présente revue narrative, qui intègre des données issues de différents domaines des sciences de la vie.
Après une première partie réaffirmant que le modèle de Sutherland n’est plus compatible avec les connaissances scientifiques actuelles, nous étudions les rôles poententiels de la vasomotion, du système nerveux autonome, de la microcirculation, du système lymphatique, de la matrice extracellulaire, du réseau fascial et des phénomènes d’entrainement dans la genèse du MRP/IRC. Sur cette base, nous proposons un modèle intégratif et soulignons la nécessité d’une évolution conceptuelle.
II. Méthodes
Cette revue a été menée conformément aux recommandations développées par Tricco et al. pour la réalisation d’une revue narrative suivant la déclaration PRISMA-AcR [21].
Le cadre théorique de cette revue de portée a été élaboré par une équipe de trois experts (F.M., L.S., A.G.) spécialisés en recherche scientifique (L.S., A.G.) et dans l’enseignement et la pratique clinique de l’ostéopathie (F.M., L.S.).
Le cadre présent est basé sur les résultats et conclusions de l’article de Mériaux et al. [1].
Question de recherche
Quelles sont les données scientifiques actuellement disponibles permettant de déterminer si les recherches contemporaines soutiennent ou remettent en question le modèle empirique du mécanisme respiratoire primaire (MRP) décrit par Sutherland [2][3] ?
Stratégie de recherche
Une recherche systématique de la littérature a été finalisée en octobre 2023 à partir des bases de données bibliographiques indexées MEDLINE, ScienceDirect et Cochrane Library [1]. Cette première recherche a permis d’identifier huit domaines de recherche susceptibles d’apporter des explications physiologiques complémentaires à l’IRC/MRP, au-delà des cinq piliers décrits par Sutherland : le système lymphatique, la vasomotricité, les ondes de Traube-Hering et de Mayer, la variabilité de la fréquence cardiaque, la fréquence ventilatoire, la matrice extracellulaire, les hypothèses métaboliques et l’embryologie.
Une recherche documentaire complémentaire et approfondie a ensuite été menée entre septembre 2024 et février 2025 à l’aide des bases PubMed, ScienceDirect, Ostmed.DR et Google Scholar. Les mots-clés ont été adaptés à chaque base de données : cranial rhythmic impulse ; primary respiratory mechanism ; Traube-Hering waves, Mayer waves ; cranial osteopathy ; osteopathy in the cranial field ; craniosacral therapy ; cranial osteopathic manipulative medicine ; heart rate variability ; cranial bone mobility ; central link ; vasomotion ; autonomic nervous system ; ventilation ; muscle tone ; microcirculation ; lymphatic system ; glymphatic systems ; extracellular matrix ; training ; embryological movement ; ideomotion ; intracranial pressure.
Aucune restriction n’a été appliquée concernant le type d’étude, la population étudiée, les critères de jugement ou la date de publication. Les articles ont d’abord été sélectionnés à partir de la lecture de leurs résumés afin d’en évaluer la pertinence, puis analysés dans leur intégralité. Les bibliographies des articles retenus ont également été examinées et une stratégie dite « boule de neige » (snowballing) a été utilisée afin d’identifier d’autres travaux pertinents.
Critères d’éligibilité
Afin de garantir une couverture aussi large que possible de la littérature disponible, aucune évaluation formelle de la qualité méthodologique des études incluses n’a été réalisée. Chaque article a en revanche fait l’objet d’une analyse critique de sa pertinence vis-à-vis de la question de recherche lors de réunions de discussion entre les auteurs (F.M., L.S., A.G.).
Les études sélectionnées ont été analysées dans le but d’identifier les mécanismes physiologiques susceptibles soit de soutenir, soit de remettre en question le modèle du MRP. Lorsqu’elles abordaient plusieurs thématiques, certaines études ont été intégrées dans plusieurs catégories d’analyse.
Synthèse et catégorisation des données
Au total, 157 articles ont été retenus et regroupés selon les thématiques suivantes :
- La motilité inhérente du système nerveux central et de la moelle épinière ;
- Les fluctuations du liquide céphalorachidien ;
- La motilité des membranes intracrâniennes et intrarachidiennes ;
- La mobilité articulaire des os du crâne ;
- Le mouvement involontaire du sacrum entre les os iliaques (« hypothèse du lien central ») ;
- L’IRC/MRP et la vasomotricité ;
- L’IRC/MRP et le système nerveux autonome (incluant les ondes de Traube-Hering-Mayer et la variabilité de la fréquence cardiaque) ;
- Le MRP, la microcirculation, le système lymphatique, la matrice extracellulaire et le réseau fascial ;
- Le MRP et les phénomènes d’entraînement (entrainment) ;
- Le MRP et les mouvements embryologiques.
Les articles ont été identifiés principalement selon une stratégie de type snowballing, à partir des références déjà incluses dans la revue systématique de Mériaux et al. [1], complétées par l’expertise des auteurs dans le domaine ainsi que par l’analyse des bibliographies des études sélectionnées.
Cadre d’interprétation
L’analyse de la littérature suggère que les cinq piliers décrits par Sutherland puis formalisés par Magoun pourraient correspondre à un modèle empirique qui n’est pas pleinement soutenu par les connaissances scientifiques actuelles. En revanche, plusieurs phénomènes physiologiques rythmiques — notamment la vasomotricité, la variabilité de la fréquence cardiaque, la microcirculation et les ondes de Traube-Hering-Mayer — apparaissent comme des candidats plausibles pour contribuer à l’élaboration d’un modèle explicatif davantage fondé sur les données scientifiques contemporaines.
III. Résultats : réévaluation de la théorie de Sutherland
Le modèle de Sutherland est solidement ancré dans la littérature.
Cependant, nous montrerons dans cette section que ce modèle ne s’aligne pas avec les connaissances scientifiques contemporaines.
III.1. La motilité inhérente du système nerveux central et de la moelle épinière
Depuis 1882, de nombreux anatomistes ont observé et décrit le mouvement continu et cyclique de la masse cérébrale et de la moelle, correspondant à une systole et une diastole, qui, selon Sutherland, pourrait agir comme une pompe hydraulique activant le liquide céphalo-rachidien (LCR) [12].
Par la suite, Harold Magoun [2], puis John Upledger [15], ont proposé une interprétation différente. Selon eux, ce serait la production rythmique du LCR par les plexus choroïdes qui mettrait l’encéphale en mouvement, et non l’inverse.
Selon le modèle du « pressurestat » d’Upledger, la sécrétion du LCR se ferait à une vitesse deux fois supérieure à sa résorption. Des mécanorécepteurs situés au niveau des sutures crâniennes détecteraient un seuil maximal de pression intracrânienne et, par un mécanisme de rétrocontrôle, interrompraient temporairement la sécrétion jusqu’à ce que la pression redescende. La production reprendrait alors, générant ainsi un cycle oscillatoire.
Cependant, ce modèle n’est pas compatible avec les connaissances actuelles. La production du LCR n’est ni intermittente ni rythmique [10]. Lorsque la pression intracrânienne augmente, la résorption du LCR augmente simultanément grâce à l’accroissement de la surface d’échange des villosités arachnoïdiennes au sein des sinus veineux. Cette description théorique est connue sous le nom de doctrine « statique » de Monro-Kellie [22][23].
Dans des conditions physiologiques, les volumes intracrâniens restent dans un état d’équilibre dynamique : toute augmentation du volume d’un compartiment (parenchyme cérébral, sang ou LCR) est compensée par une diminution d’un autre. Cette compensation n’est toutefois pas instantanée et génère des variations transitoires de pression intracrânienne (PIC) [24].
Ces variations de PIC sont d’une grande importance clinique et sont notamment surveillées chez les patients souffrant de traumatismes crâniens ou d’hydrocéphalie. Elles peuvent être enregistrées de manière non invasive à l’aide de capteurs mécaniques placés sur le cuir chevelu, capables de détecter les déformations micrométriques du crâne battement après battement [25–27]. Dès 1985, Pitlyk et al. avaient montré que ces faibles expansions crâniennes reflétaient les variations de pression intracrânienne [28].
Dans le domaine temporel, le signal de PIC est constitué de plusieurs composantes superposées. L’analyse fréquentielle permet toutefois de distinguer :
- les ondes lentes (0,3 à 3 cycles par minute) ;
- les ondes respiratoires (8 à 20 cycles par minute) ;
- les ondes cardiaques (40 à 160 cycles par minute) [29][24].
Vinje et al. [30] ont mesuré les gradients de pression intracrânienne à l’aide de deux capteurs implantés simultanément. Leur analyse spectrale a mis en évidence deux pics principaux : l’un correspondant au rythme artériel (0,7 à 1,6 Hz ; 42 à 96 battements/minute), l’autre à l’activité respiratoire (0,15 à 0,4 Hz ; 9 à 24 respirations/minute). Fait intéressant, ils ont également observé des oscillations de basse fréquence inférieures à 0,1 Hz (environ 6 cycles/minute), sans toutefois évaluer leur contribution au flux du LCR [30].
En utilisant l’imagerie par résonance magnétique (IRM), Maier et al. [13] ont déjà observé un mouvement du cerveau et du LCR qui semblait lié à l’activité cardiaque et respiratoire.
La masse cérébrale et la moelle se déplaçaient caudalement et médialement (2-3 mm) pendant la systole ; l’inverse se produisait pendant la diastole [11][14].
Lors d’une inspiration, le système nerveux est poussé vers le crâne et le cerveau se rétracte, tandis que lors d’une expiration, le mouvement prédominant est caudal et le cerveau s’expanse [31].
Ces observations montrent que les mouvements du névraxe sont principalement gouvernés par la dynamique cardio-respiratoire, à une fréquence proche de celle de la respiration [32], donc plus rapide que celle habituellement attribuée au MRP/CRI.
Dans l’ensemble, ces données suggèrent que le MRP/CRI n’est pas directement lié à une motilité intrinsèque du cerveau ni au rythme respiratoire.
III.2. Fluctuation du liquide céphalo-rachidien (LCR)
Le LCR est un liquide biologique transparent entourant le cerveau et la moelle épinière, entre la pie-mère et l’arachnoïde, dans l’espace sous-arachnoïdien.
Il a un rôle de protection, et transporte les nutriments et les déchets.
Le LCR est principalement sécrété par filtration du sang au niveau du plexus choroïde des ventricules cérébraux [33].
En tant que liquide, le LCR est peu compressible.
Ainsi, il pourrait pousser les membranes qui feraient bouger les os du crâne.
La théorie initiale était que la circulation pulsatile rythmique du LCR pourrait être à l’origine de l’IRC/MRP.
Le flux du LCR est principalement entraîné par le cycle cardiaque et est donc pulsatile.
Les mouvements du LCR sont également influencés par les mouvements respiratoires, la posture, la pression des veines jugulaires, la pression artérielle et l’activité physique [34][35][30].
Les mouvements de la paroi artérielle constituent le principal moteur du déplacement du LCR, par un mécanisme appelé « pompage périvasculaire » (perivascular pumping). Ainsi, la vitesse du LCR suit étroitement celle des mouvements artériels.
Le volume total de LCR est d’environ 150 mL et se renouvelle trois à quatre fois par jour, soit près de 500 mL produits quotidiennement [36]. Le LCR circule depuis les ventricules vers l’espace sous-arachnoïdien avant d’être résorbé principalement par les voies veineuses et lymphatiques.
Les données actuelles indiquent que le système veineux assure environ 30 % du drainage du LCR, tandis que le système lymphatique nasal représente une voie majeure, participant à environ 40 % du drainage via les nerfs crâniens, notamment au niveau de la lame criblée de l’ethmoïde [37]. Environ 25 % du drainage se fait au niveau rachidien via les villosités arachnoïdiennes associées aux veines spinales. Une faible proportion diffuse également à travers l’arachnoïde et la dure-mère vers les tissus conjonctifs environnants avant de rejoindre les réseaux veineux et lymphatiques [37][38].
Sutherland postulait une continuité entre le LCR et l’ensemble de l’organisme via des voies lymphatiques, périvasculaires et périneurales, ce qui permettrait selon lui de percevoir ce rythme partout dans le corps [2]. Les données anatomiques actuelles contredisent cependant cette hypothèse. Sakka et al. [38] ont montré que l’espace sous-arachnoïdien se rétrécit à proximité des ganglions nerveux et se termine en cul-de-sac anatomique, sans continuité directe avec les tissus périphériques.
La continuité s’effectue plutôt via les espaces périvasculaires de Virchow-Robin, qui semblent davantage impliqués dans le drainage du liquide interstitiel cérébral que dans celui du LCR lui-même. De plus, les trabéculations et ligaments arachnoïdiens ralentissent et perturbent sa circulation, laquelle demeure lente (environ 1 cm/h), non linéaire et irrégulière.
Les progrès récents ont néanmoins montré que le LCR pouvait diffuser vers le système nerveux périphérique. Ligocki et al. [39] ont démontré que des nanoparticules injectées dans le LCR pouvaient migrer du système nerveux central vers le système nerveux périphérique jusqu’au périnèvre, à l’endonèvre et même à l’axoplasme des nerfs distaux. De même, Pessa et al. [40] ont décrit des circulations du LCR au sein des gaines nerveuses et des voies glymphatiques, suggérant l’existence d’un système circulatoire ouvert dans les tissus nerveux.
Il n’existe cependant aucune preuve d’un flux continu, palpable et rythmé du LCR tel qu’il est traditionnellement décrit en ostéopathie. Ni sa production ni son drainage ne sont des phénomènes rythmiques.
Au sein du système cranio-spinal, la dynamique du LCR est fortement couplée aux variations de pression intrathoracique induites par la respiration. L’inspiration favorise un déplacement crânial du LCR, tandis que l’expiration induit un déplacement caudal [41][32]. Ces phénomènes témoignent de l’étroite interaction entre le LCR et la circulation veineuse.
La vitesse du flux varie selon les régions anatomiques. Elle est plus faible dans les zones éloignées des sites de production et des pulsations artérielles, et devient minime voire absente au niveau lombaire [36].
En conclusion, les mouvements d’expansion et de rétraction du cerveau semblent principalement liés à la respiration [30] plutôt qu’aux variations de pression du LCR, dont l’amplitude est trop faible pour entraîner à elle seule des mouvements crâniens significatifs [10]. De plus, aucune continuité anatomique n’existe entre le réservoir de LCR et les tissus périphériques. Il paraît donc peu probable que la dynamique du LCR, à elle seule, explique le MRP/IRC perçu par les ostéopathes. Bien que le LCR occupe une place centrale dans la philosophie de l’ostéopathie crânienne et que l’amélioration de ses fluctuations puisse participer à certains effets thérapeutiques, rien n’indique qu’il constitue l’origine du rythme étudié ici.
III.3. Motilité des membranes intracrâniennes et intraspinales
La dure-mère fait partie du système méningé, un ensemble d’enveloppes membraneuses entourant l’encéphale et le système nerveux central. Au niveau intracrânien, elle s’attache à certains os du crâne et peut se prolonger à l’extérieur à travers les sutures crâniennes [42]. Elle présente également des attaches sur les vertèbres cervicales supérieures et sur le sacrum, ainsi que tout au long du rachis [43][44][45].
À l’intérieur du crâne, la faux du cerveau (falx cerebri) et la tente du cervelet (tentorium cerebelli) font partie du système dural et participent à une structure reliant mécaniquement les différents os du crâne. Sutherland désignait cet ensemble sous le terme de « membrane de tension réciproque » (reciprocal tension membrane). Selon lui, les variations de tension de la dure-mère contribuaient aux déplacements des os auxquels elle est attachée et un tel mécanisme réciproque ne pouvait exister que si cette membrane demeurait constamment tendue [3].
Dans ce modèle, le crâne effectuerait des mouvements rythmiques au cours de phases dites d’inspiration et d’expiration, sous l’effet des variations de tension au sein de ces membranes de tension réciproque. Pourtant, la dure-mère possède une épaisseur moyenne d’environ un millimètre et présente des propriétés viscoélastiques anisotropes. Elle n’est donc pas inextensible, contrairement à ce qu’ont pu suggérer certaines théories ostéopathiques [46].
Les travaux d’Al-Habib et al. [47] confirment les connaissances anatomiques fondamentales : parmi les tissus étudiés, la dure-mère est la plus élastique, suivie par la pie-mère puis par la moelle épinière. Leur étude montre également qu’une moelle comprimée devient significativement plus rigide qu’une moelle non comprimée [47].
Ces résultats rejoignent ceux de Royo-Salvador et al. [48][49] concernant le conflit de croissance cranio-cervical. Selon leurs travaux, une mise en tension du filum terminale pourrait contribuer à une traction et une compression de la moelle épinière, entraînant secondairement une descente du cervelet vers le foramen magnum. La section du filum terminale a notamment été associée à une ascension du cône médullaire [48][49].
Les méninges transmettent les contraintes mécaniques de l’extérieur vers l’intérieur, comme lors d’un traumatisme, mais également de l’intérieur vers l’extérieur [3]. Elles reflètent aussi les conditions mécaniques et métaboliques de leur environnement. Au fil du temps, elles peuvent modifier à la fois leur capacité à répondre aux contraintes et leur structure intrinsèque [50].
Selon Bordoni et al. [50], les méninges sont donc capables de s’adapter aux tensions et aux sollicitations mécaniques. Elles ne semblent pas suffisamment rigides pour déplacer les os du crâne, sauf dans certaines situations pathologiques ou liées à l’âge où elles peuvent s’ossifier, notamment au niveau des sinus veineux duraux. Une telle ossification pourrait alors modifier la transmission des contraintes mécaniques [50].
À la lumière des données actuelles, si une force de tension — qu’il s’agisse d’une pression ou d’une traction — était transmise depuis l’encéphale vers la peau, les différentes couches tissulaires intermédiaires devraient en amortir l’amplitude et ralentir sa propagation [51]. Bordoni [50] suggère également que les propriétés viscoélastiques des fascias atténueraient toute onde se propageant du crâne vers la périphérie, tout comme elles amortiraient une onde se déplaçant de l’intérieur du crâne vers l’extérieur.
Par conséquent, si l’IRC/MRP était transmis mécaniquement depuis la région crânienne, son amplitude devrait diminuer progressivement à mesure que l’on s’éloigne de l’axe vertébral. Or, les ostéopathes qui utilisent ce concept décrivent généralement une fréquence identique, voire synchronisée, dans différentes régions du corps. Cette observation rend donc peu probable l’hypothèse d’une transmission purement mécanique.
III.4. Mobilité articulaire des os du crâne
La première hypothèse formulée par Sutherland en 1939 était que le MRP perçu au niveau crânien résultait d’un mouvement des os du crâne. Dans ce modèle, les os étaient supposés être mobilisés par les membranes, elles-mêmes influencées par les fluctuations du liquide céphalorachidien. Sutherland considérait que le crâne conservait, même chez l’adulte, une certaine souplesse au niveau des sutures.
Historiquement, la mobilité des os du crâne était considérée comme une impossibilité anatomique. Elle demeure aujourd’hui l’un des aspects les plus controversés du MRP [52].
L’argument principal avancé par les défenseurs de cette théorie est que les sutures reliant les différentes parties du crâne ne s’ossifient jamais complètement [53][54]. Bien que leur fermeture soit progressive, des ponts osseux apparaissent bien avant la fusion complète [54]. La rigidité du crâne augmente avec l’âge, mais avec une grande variabilité selon les individus et selon les sutures concernées [55].
La calcification de la majorité des sutures débute vers 20 à 30 ans [56], mais l’achèvement de ce processus varie considérablement selon les individus et les types de sutures [57][58][59]. Après 60 ans, la plupart des sutures sont totalement ossifiées [60].
Pourtant, certains ostéopathes ne rapportent aucune difficulté particulière à percevoir ou à restaurer l’IRC/MRP chez les sujets âgés [61]. Si le MRP dépendait directement de la mobilité suturale, son amplitude et sa force devraient diminuer progressivement avec l’ossification des sutures au cours du vieillissement. Cela ne semble pas être observé de façon constante.
Dans le modèle ostéopathique crânien, la synchondrose sphéno-basilaire est considérée comme la structure centrale de la biomécanique crânienne. C’est également à cette articulation que Sutherland attribuait plusieurs dysfonctions crâniennes [2][3].
Or, la synchondrose sphéno-basilaire commence à s’ossifier avant la puberté et ce processus s’achève au cours de celle-ci [46]. Plus généralement, les sutures du chondrocrâne humain fusionnent progressivement au cours de l’enfance et de l’adolescence, jusqu’à la fin de la croissance crânienne [60][62][63].
Chez l’adulte, la synchondrose sphéno-basilaire est donc ossifiée. D’un point de vue scientifique, elle ne peut plus être considérée comme le moteur principal des mouvements crâniens ni comme l’origine des dysfonctions décrites en ostéopathie crânienne. Les explications justifiant les approches manuelles visant à « libérer » cette articulation devraient par conséquent être réévaluées [46].
Certains auteurs ont tenté de déterminer si les faibles pressions habituellement appliquées par les praticiens (5 à 10 g) étaient capables de mobiliser les sutures. Downey et al. [64] ont ainsi émis l’hypothèse que des charges légères appliquées sur l’os frontal de lapins anesthésiés, reproduisant la technique ostéopathique du frontal lift, pouvaient induire un déplacement mesurable de la suture coronale. Ils n’ont cependant pas pu démontrer cet effet.
Pour obtenir un déplacement de 1 mm d’une suture présentant un degré d’ossification comparable à celui d’un adulte de 20 à 30 ans, une force d’environ 50 kg serait nécessaire chez le lapin [65]. Chez un enfant, cette force est estimée à environ 15 kg, soit très largement au-dessus des 5 à 10 g généralement recommandés en ostéopathie crânienne [66].
Pendant longtemps, les scientifiques ont considéré les os du crâne comme fusionnés et immobiles. Nous savons aujourd’hui que le crâne possède néanmoins une certaine flexibilité, comme la plupart des tissus de l’organisme [67][55]. Le tissu osseux possède en effet des propriétés viscoélastiques liées à sa composition en collagène et en élastine, lui permettant des déformations mécaniques de l’ordre de 10 à 15 % [46].
Starkey [61] décrit notamment le clivus sphénoïdal comme une structure spongieuse, donc potentiellement déformable, mais également épaisse et résistante. Cook [68] parle davantage de flexibilité que de mobilité des os crâniens, tandis que Seimetz et al. [55] évoquent une possible motilité des os du crâne.
Bien que l’os soit l’un des tissus les plus rigides du corps, il se déforme lors du fonctionnement normal et plus encore lors de traumatismes. Plusieurs études ont montré que des contraintes internes ou externes pouvaient modifier la forme du crâne chez l’animal comme chez l’humain.
À l’aide de l’IRM, Crow et al. [69] ont observé des variations significatives de plusieurs mesures crâniennes sans intervention humaine particulière. Des différences significatives ont été retrouvées pour la surface, la largeur, la hauteur et le grand axe du crâne, mais pas pour son périmètre ni son petit axe. Ces résultats ne correspondent pas réellement aux mouvements d’expansion et de rétraction décrits par les ostéopathes.
Le modèle crânien de Sutherland accorde relativement peu de place à l’action des muscles. Cela s’explique probablement par le fait qu’il travaillait essentiellement sur des os secs, comme c’était souvent le cas dans les premiers modèles ostéopathiques. Pourtant, selon Gabutti et al. [70], de nombreuses recherches consacrées aux propriétés mécaniques des os du crâne et des sutures montrent que la contraction musculaire constitue l’une des principales causes de déformation des os crâniens [71].
En raison de leurs propriétés viscoélastiques et de la composition de leur matrice extracellulaire environnante — riche en fibres de collagène, en protéoglycanes et en eau — les sutures crâniennes sont nettement plus souples que l’os adjacent [70]. Plus qu’un rôle de mobilité, elles semblent assurer des fonctions d’absorption des contraintes et de transmission des forces [72][73].
Les propriétés biomécaniques du crâne humain et de ses sutures varient selon l’âge ainsi que selon les différentes couches anatomiques concernées, notamment le diploë et les tables interne et externe [74]. Les sutures peuvent influencer la manière dont le crâne se déforme, mais cette déformation persiste même lorsque les sutures sont fusionnées. La densité suturale, qui dépend du degré de fermeture des sutures, pourrait néanmoins limiter l’amplitude de ces déformations [70].
En conclusion, les os du crâne présentent bien des mouvements, mais pas nécessairement les uns par rapport aux autres. Dans les travaux d’Adams et de Heisey, l’amplitude des déplacements mesurés au niveau de la suture sagittale était de l’ordre de 300 µm [75][52].
La mobilité crânienne serait ainsi davantage une déformabilité globale du crâne qu’une mobilité articulaire entre ses différentes pièces osseuses. Cette déformabilité pourrait résulter des variations de pression intracrânienne, des fluctuations du liquide céphalorachidien, des tensions membranaires, mais également de forces externes telles que les contractions des muscles crâniens et des tissus conjonctifs associés. Les vaisseaux sanguins traversant ces tissus pourraient également contribuer à ces déformations par leur propre motilité [76].
À l’aide d’examens IRM sériés, Moskalenko et al. [76] ont mesuré des variations rythmiques de la forme et du volume du crâne. Ils ont confirmé l’existence d’un rythme compris entre 6 et 15 cycles par minute ainsi que l’extensibilité physique de la boîte crânienne. Ils ont également observé une augmentation du diamètre crânien de 0,2 à 0,4 mm immédiatement après l’injection de 20 mL de liquide dans l’artère carotide, démontrant ainsi qu’une augmentation du volume intracrânien peut effectivement modifier la forme du crâne [76].
La mobilité crânienne devrait donc être considérée davantage comme une conséquence que comme une composante du MRP : un effet plutôt qu’une cause. Cette observation invite à reconsidérer le modèle proposé par Sutherland.
Du point de vue thérapeutique, cela n’exclut pas pour autant la possibilité de diminuer les tensions présentes au niveau d’une suture crânienne, comme on le ferait pour n’importe quelle autre structure fasciale, quel que soit son degré d’ossification. Hamm [77] suggère notamment qu’une densification locale des tissus, liée à une polymérisation des fibres de collagène, pourrait rigidifier les berges d’une suture et entraîner une forme de fixation.
Dans cette perspective, le traitement ostéopathique crânien viserait davantage à assouplir les tissus de la région concernée, comme ailleurs dans le corps, qu’à restaurer un mouvement entre les os du crâne eux-mêmes.
III.5. Le mouvement involontaire du sacrum entre les os iliaques. L’hypothèse du « core link »
Sutherland considérait la dure-mère comme une structure inextensible. Dans ce modèle, la traction générée par les mouvements des os du crâne serait transmise par la dure-mère et entraînerait le mouvement du sacrum [2][3]. Cependant, si la dure-mère était réellement rigide, il serait difficile d’expliquer la possibilité même de la flexion du rachis.
En réalité, la dure-mère possède des propriétés élastiques. Des études expérimentales montrent que, lorsque la colonne vertébrale est en position neutre, la gaine durale spinale présente des replis, suggérant que le tissu est relâché plutôt que continuellement tendu [78]. Al-Habib et al. [47] ont montré que la dure-mère possède une élasticité environ deux fois supérieure à celle de la pie-mère et trois fois supérieure à celle de la moelle épinière. Par ailleurs, la longueur du canal rachidien varie de 19,4 ± 6,4 mm entre la flexion et l’extension de la colonne vertébrale [79]. Ces données ne soutiennent donc pas l’hypothèse d’un lien dural inextensible capable d’entraîner mécaniquement les mouvements du sacrum.
Cella et al. [80] ont étudié la relation entre l’occiput et le sacrum sous un angle neurophysiologique. Une technique appliquée au sacrum n’a entraîné aucune modification immédiate de l’activité cérébrale occipitale dans la bande alpha. À l’inverse, la technique crânienne dite de « compression du quatrième ventricule » (CV4), conformément à des travaux antérieurs [81], a produit des effets immédiats. Ces résultats suggèrent que la relation ostéopathique entre le crâne et le sacrum pourrait reposer sur un mécanisme biologique différent d’une simple transmission mécanique par la dure-mère.
Ce pilier du modèle de Sutherland a également donné naissance à l’idée d’une unité fonctionnelle et d’une synchronisation des fréquences entre le crâne et le bassin, puis par extension dans l’ensemble du corps. Toutefois, cette conception ne fait pas consensus parmi les ostéopathes. Moran et Gibbons [82] n’ont pas réussi à démontrer une synchronisation entre les rythmes palpés au niveau du crâne et du sacrum. De même, Rogers et al. [83] n’ont observé aucune correspondance entre les rythmes perçus au niveau de la tête et du pied lorsque deux examinateurs palpitaient simultanément le MRP.
On pourrait objecter que les résultats auraient pu être différents si les sujets avaient préalablement bénéficié d’un traitement ostéopathique visant à les « rééquilibrer ». Des études expérimentales complémentaires, réalisées en aveugle, seraient utiles pour mesurer le MRP en différents points du corps avant et après un traitement ostéopathique, en faisant intervenir un même praticien ou plusieurs praticiens.
À la lumière des travaux d’Al-Habib et al. [47] et de Royo-Salvador et al. [48][49], si un mouvement était réellement transmis mécaniquement du crâne vers le sacrum, il devrait emprunter la moelle épinière et le filum terminale. Or, le filum terminale s’insère sur le coccyx et non sur le sacrum [47][48][49]. Cette observation ne correspond pas au modèle proposé par Sutherland [2][3].
En conclusion de cette première partie, nous avons montré que le modèle physiologique du MRP/IRC proposé par Sutherland en 1939, bien qu’encore largement enseigné comme un dogme, est aujourd’hui remis en question par les connaissances scientifiques contemporaines. Nous allons maintenant examiner différentes hypothèses et modèles explicatifs alternatifs susceptibles d’être confrontés aux données actuelles, dans la perspective de faire émerger un futur modèle consensuel.
IV. IRC/MRP et vasomotricité
Le MRP est décrit comme un mouvement cyclique perceptible sous les mains dans l’ensemble du corps et pas uniquement au niveau du crâne, où il est généralement désigné sous le terme d’IRC. Dès 1850, Jones [84] mit en évidence l’existence d’une rythmicité vasculaire à la face ventrale de l’aile de chauve-souris. Ce rythme était indépendant des rythmes cardiaque et respiratoire et correspondait à des variations spontanées et rythmiques du diamètre des veines. Ces variations résultaient de contractions intermittentes des muscles lisses présents dans la paroi des microvaisseaux, produisant un déplacement actif du sang sous forme d’oscillations lentes. Jones désigna ce phénomène sous le terme de flowmotion, aujourd’hui plus communément appelé vasomotion ou vasomotricité.
La vasomotricité correspond à la capacité des vaisseaux sanguins ou lymphatiques à se contracter et à se dilater en réponse à des variations de pression, modulant ainsi le débit circulatoire [85]. Au niveau artériel, les gros vaisseaux, comme l’aorte et les artères de gros calibre, répondent principalement de manière passive en raison de l’abondance de fibres élastiques dans leur paroi. Cette propriété, appelée compliance ou distensibilité artérielle, permet d’amortir les effets de la systole ventriculaire sur la pression artérielle [86]. Durant la diastole, ces gros troncs artériels retrouvent leur diamètre initial à la manière d’un ballon qui se dégonfle, limitant ainsi la chute de la pression artérielle [86].
Deux autres mécanismes interviennent spécifiquement au niveau des petites artères de résistance et des artérioles, dont la média est riche en cellules musculaires lisses vasculaires (vascular smooth muscle cells, VSMC). Ces mécanismes sont absents des capillaires et des gros troncs artériels, dépourvus de telles structures musculaires. Le premier mécanisme est dit myogène, le second neurogène. Une certaine confusion persiste parfois dans la littérature lorsque les auteurs ne précisent pas clairement lequel de ces mécanismes est étudié.
La vasomotricité occupe une place importante dans l’étude des troubles vasculaires. À l’aide de la spectroscopie proche infrarouge (NIRS), Fredriksson et al. [87] ont mesuré des oscillations hémodynamiques spontanées faibles et très faibles (0,003–0,15 Hz), confirmant ainsi les observations antérieures de Stefanovska et al. [88]. Ces derniers ont décrit plusieurs bandes fréquentielles correspondant à différents mécanismes physiologiques : endothéliale (0,0095–0,02 Hz), neurogène (0,02–0,06 Hz), myogène (0,06–0,15 Hz), respiratoire (0,15–0,4 Hz) et cardiaque (0,4–1,6 Hz) [88].
Le mécanisme myogène est celui qui est le plus souvent étudié dans les travaux consacrés à la vasomotion. Les artères de petit diamètre se contractent en réponse à une augmentation brutale de la pression artérielle et se relâchent lorsque cette pression diminue [89][90]. Au niveau des bifurcations artériolaires, certaines cellules présentent une activité de type pacemaker générée par des oscillations spontanées de la concentration intracellulaire en calcium (Ca²⁺) au sein des cellules musculaires lisses vasculaires [91][90][92].
Ces cellules répondent également à différents signaux : signaux adrénergiques vasoconstricteurs, signaux cholinergiques vasodilatateurs, facteurs métaboliques (pH, CO₂, ATP, NO, O₂), facteurs humoraux (prostaglandines, EDRF, etc.) ainsi qu’à des influences myogènes telles que la température ou les pressions musculaires locales [90].
Dans leur étude consacrée à la vasomotion artériolaire et à la régulation du débit sanguin, Colantuoni et al. [93] ont identifié la variation du diamètre vasculaire au cours du temps comme l’une des principales caractéristiques de la microvascularisation artériolaire. L’analyse spectrale a montré que la fréquence fondamentale variait approximativement de 4 à 15 cycles par minute dans les artérioles de rang 1 (8 µm) et de 2 à 11 cycles par minute dans les artérioles de rang 2 (10–13 µm). Les vaisseaux de troisième et quatrième ordre (20 et 30 µm respectivement) présentaient des fréquences plus basses, comprises entre 0,5 et 6 cycles par minute et entre 0,3 et 3 cycles par minute [93].
Ces oscillations sont synchronisées par des phénomènes électriques liés aux variations du potentiel membranaire cellulaire, c’est-à-dire aux processus de polarisation et de dépolarisation. Cette activité est transmise par les jonctions communicantes (gap junctions) aux cellules voisines, y compris à celles ne possédant pas elles-mêmes d’activité pacemaker [94][95].
La conduction longitudinale des réponses vasomotrices permet la coordination des modifications de diamètre et de la distribution des flux au sein du réseau vasculaire. Elle participe également à la régulation de la résistance vasculaire en assurant l’intégration fonctionnelle des segments vasculaires proximaux et distaux au sein de la microcirculation [96].
Les influences neurogènes ne sont pas indispensables à ce mécanisme, bien qu’une stimulation adrénergique puisse le renforcer. Cette variation n’est donc pas nécessairement rythmique, cyclique ou synchronisée avec les vaisseaux voisins. Il s’agit plutôt d’un phénomène localisé et intermittent, ce qui rend son association directe avec le MRP difficile à établir.
Nilsson et Aalkjaer [91] soulignent d’ailleurs que : « Dans de nombreux contextes expérimentaux, la vasomotricité pose problème ; par exemple, il est difficile de définir précisément le tonus d’un vaisseau lorsqu’il oscille. De plus, la vasomotricité est fréquemment imprévisible et difficile à reproduire ; il arrive que les animaux de laboratoire cessent de présenter une activité vasomotrice pendant des périodes variables, conduisant parfois à l’échec des expériences in vivo lorsque ce phénomène ne peut être reproduit. » Hamm [77] arrive à une conclusion similaire : « La présence, tout comme l’intensité ou l’importance de la vasomotricité, est extrêmement variable. »
La vasomotricité n’est d’ailleurs pas systématiquement observée dans des conditions physiologiques normales [97]. Elle apparaît plus fréquemment lorsque la perfusion tissulaire est réduite, est souvent déclenchée par un stress métabolique et demeure généralement absente au repos [94]. Elle peut également ne survenir que de manière transitoire [77]. Ainsi, la vasomotricité d’origine myogène pourrait participer au mécanisme du MRP/IRC, sans toutefois pouvoir en constituer l’unique explication. Si elle joue effectivement un rôle, son caractère imprévisible pourrait contribuer à expliquer les faibles niveaux de reproductibilité intra- et inter-examinateurs rapportés dans les études palpatoires.
Cependant, Nilsson et Aalkjaer [91] ont également observé des vasoconstrictions rythmiques, probablement d’origine neurogène. Les artérioles transversales de plus grand calibre présentaient les fréquences les plus basses, comprises entre 0,3 et 3 cycles par minute, avec des variations de diamètre représentant 5 à 20 % du diamètre moyen [91]. Les artérioles d’environ 18 μm de diamètre présentaient des amplitudes de variation de 15 à 50 % et des fréquences comprises entre 0,5 et 6 cycles par minute. Pour les artérioles d’environ 11 μm, les amplitudes atteignaient 50 à 100 % du diamètre moyen, avec des fréquences comprises entre 2 et 11 cycles par minute. Enfin, les plus petites artérioles terminales, les plus superficielles, présentaient les fréquences les plus élevées, comprises entre 4 et 15 cycles par minute, ainsi que les amplitudes les plus importantes, variant de 60 à 100 % de leur diamètre moyen.
Fait particulièrement intéressant, dans leur étude réalisée chez le hamster, Nilsson et Aalkjaer [91] rapportent une prédominance d’oscillations à 0,1 Hz. Ces ondes étaient transmises à partir des artérioles de taille intermédiaire (type 3 selon la classification de Strahler) et se propageaient vers les artérioles plus petites (types 2 et 1), soit une fréquence très proche de celle généralement attribuée au MRP. Cette observation suggère une superposition de différents rythmes artériolaires, les oscillations les plus superficielles, générées par les plus petites artérioles, étant probablement les plus facilement perceptibles sous les mains de l’ostéopathe.
Villermain-Lecolier et al. [98][99] ont décrit l’existence de micro-mouvements périodiques perceptibles à la surface de la peau, d’une amplitude comprise entre 20 et 50 μm. Ces oscillations présentaient des fréquences distinctes des rythmes cardiaque et respiratoire, également enregistrés chez les mêmes sujets. Leur fréquence moyenne était de 9,73 ± 3,48 cycles par minute. Selon ces auteurs, le MRP serait l’expression d’une vasomotricité locale et ne nécessiterait pas d’être synchronisé à l’échelle de l’ensemble du corps. Comme nous l’avons déjà souligné, il n’existe toutefois pas de consensus clair parmi les ostéopathes sur ce point.
La vasomotricité favorise la circulation sanguine et la perfusion tissulaire, contribuant ainsi à la vitalité de l’organisme. L’exploration du lien entre MRP/IRC et vasomotricité s’inscrit pleinement dans l’un des principes fondamentaux de l’ostéopathie : l’importance de la circulation dans le maintien de la santé, telle que soulignée par Still [100].
La vasomotricité pourrait donc participer aux mécanismes physiologiques sous-jacents au MRP/IRC, mais elle ne semble vraisemblablement pas en constituer l’unique explication.
V. IRC/MRP et système nerveux autonome
V.1. IRC/MRP et ondes vasomotrices
Les ondes vasomotrices, également appelées ondes de Traube–Hering–Mayer (THM), sont des oscillations physiologiques générées par les pulsations spontanées des vaisseaux artériels, veineux et lymphatiques. Les ondes de Traube–Hering résultent de l’interaction entre l’arythmie sinusale respiratoire et la modulation de la pression pulsée chez les sujets sains [101]. Les ondes THM sont médiées par le système nerveux autonome et, associées à une variabilité élevée de la fréquence cardiaque, sont considérées comme des marqueurs d’un bon équilibre neurovégétatif [36].
Les mécanismes neurogènes influençant la vasomotricité sont présents dans des conditions physiologiques normales. Ils génèrent des contractions rythmiques et synchrones activées par le système nerveux sympathique (SNS), à l’origine d’oscillations de la pression artérielle appelées ondes de Mayer [102]. Ces ondes constituent également des réponses baroréflexes aux variations de pression artérielle [103] et seraient le résultat d’un phénomène de résonance au sein de la boucle baroréflexe sympathique [104].
Les signaux afférents provenant des artères de conductance initieraient une boucle de rétrocontrôle baroréflexe. Ces informations sont relayées par le noyau du tractus solitaire (nucleus tractus solitarius, NTS), puis transmises aux artérioles via les fibres sympathiques vasoconstrictrices de la chaîne ganglionnaire, déclenchant ainsi la vasomotricité associée aux ondes de Mayer [91].
Le NTS est situé dans le plancher du quatrième ventricule. Il est intéressant de noter que cette localisation correspond anatomiquement à la région ciblée par la technique crânienne dite de « compression du quatrième ventricule » (CV4), largement utilisée en ostéopathie crânienne pour stimuler ou restaurer le MRP.
Dans les circulations régionales, les ondes de Mayer se propagent aux artères de conductance [102]. Bien qu’atténuées par la compliance vasculaire locale, elles demeurent détectables dans certains territoires périphériques, notamment au niveau des doigts, où elles peuvent être enregistrées par photopléthysmographie (PPG) [105].
La bande fréquentielle des ondes de Mayer (0,04–0,15 Hz) correspond à la composante basse fréquence (LF) de la variabilité de la fréquence cardiaque. Cette composante est associée aux stimulations provenant des barorécepteurs et des chémorécepteurs du sinus carotidien, transmises par le système nerveux sympathique aux cellules musculaires lisses vasculaires (VSMC). Elle se distingue de la composante haute fréquence (HF), correspondant à l’arythmie sinusale respiratoire, également appelée bande des ondes de Traube–Hering (TH), synchronisée avec les mouvements respiratoires (0,15–0,4 Hz) [106].
Les ondes de Mayer présentent une cohérence forte et significative avec l’activité nerveuse sympathique efférente, c’est-à-dire une forte relation linéaire entre les fluctuations de ces variables dans le domaine fréquentiel [107]. Au sein d’une même espèce, leur fréquence est relativement stable. Chez l’être humain, elle ne semble dépendre ni du sexe, ni de l’âge, ni de la posture [107]. Elle dépend principalement du délai séparant une modification de l’activité sympathique de la réponse vasculaire qui en découle.
Ce délai est lui-même lié à la longueur des neurones sympathiques post-ganglionnaires, notamment des fibres A myélinisées à conduction lente, et donc, en définitive, à la taille de l’animal. Plus l’animal est grand, plus les fibres sont longues, plus le délai est important et plus la fréquence est basse. Ainsi, la fréquence des ondes de Mayer est d’environ 0,1 Hz chez l’Homme, 0,3 Hz chez le lapin et 0,4 Hz chez le rat [108][102]. Une observation similaire est réalisée en ostéopathie animale concernant la fréquence du MRP : plus l’animal est grand, plus la fréquence perçue est faible.
Fernandez et Lecine semblent avoir été les premiers à enregistrer les ondes vasomotrices tout en les comparant simultanément à la palpation crânienne [109].
Nelson et al. [17] ont également mis en évidence une forte corrélation entre la palpation de l’IRC et l’enregistrement des ondes THM par vélocimétrie Doppler laser (328 paires de données ; corrélation = 1,00 ; significativité = 0,00). Cette technique mesure la vitesse du flux sanguin en détectant les variations de déplacement des érythrocytes, principalement via l’hémoglobine, dans les capillaires sous-cutanés [110].
Toutefois, comme d’autres auteurs, ils ne distinguaient pas clairement les ondes de Traube–Hering, les ondes de Mayer et les oscillations THM combinées, ce qui complique l’interprétation de leurs résultats. Ils ont néanmoins conclu que les oscillations THM et le MRP/IRC étaient simultanés, voire qu’ils correspondaient à un seul et même phénomène.
Ces travaux ont ouvert de nouvelles perspectives quant à l’interprétation des fondements théoriques du MRP/IRC et des thérapies crâniennes. Nelson et al. ont attribué le phénomène de still point à une interruption transitoire du rythme du MRP, observée dans 79 % des cas et associée à une diminution de l’amplitude des ondes TH. Hamm [77] rapporte que McGrath, dans son analyse de l’article de Ferguson, proposait une interprétation similaire en suggérant que l’IRC pourrait être « la manifestation d’un phénomène extracrânien lié au flux sanguin ».
Christ et al. [111] ont mesuré des variations cycliques du volume des membres dans la gamme de fréquence du MRP/IRC et ont suggéré que ces variations étaient liées aux fluctuations de pression artérielle, à la vasomotricité artériolaire et possiblement aux variations de diamètre des vaisseaux lymphatiques. Ils ont observé une relation entre les augmentations cycliques du volume artériel et les modifications du volume des membres (< 0,177 mL pour 100 mL de tissu).
Les muscles étant pratiquement incompressibles [112] et les tissus environnants ne comportant pas d’espaces vides susceptibles d’absorber ces variations de volume, les dilatations et contractions cycliques des gaines fasciales induites par les variations du volume vasculaire, qu’il soit intra- ou extramusculaire, pourraient influencer l’organisation hélicoïdale croisée des fibres présentes dans leurs parois [113]. Si ce mécanisme est effectivement impliqué, il pourrait contribuer à expliquer les sensations de rotation interne et externe décrites par les ostéopathes lors de la perception du MRP au niveau des membres.
Au niveau crânien, Abenavoli et al. [114][115] ont conclu que les ostéopathes étaient capables de percevoir des modifications de l’amplitude de l’IRC après réalisation du test de Queckenstedt, qui consiste en une compression bilatérale des veines jugulaires internes entraînant une augmentation de la pression intracrânienne. Ces résultats suggèrent que les variations de débit et de pression veineux et artériels pourraient participer à l’origine et à la perception du MRP/IRC.
Les ondes de Mayer sont abolies, ou du moins fortement atténuées, par le blocage pharmacologique des récepteurs alpha-adrénergiques [102]. Cela suggère que leur origine hémodynamique repose principalement sur des oscillations du tonus vasomoteur sympathique des vaisseaux artériels. À l’inverse, le blocage aigu des récepteurs bêta-adrénergiques a peu ou pas d’effet sur ces oscillations [108][116], ce qui indique que le contrôle vagal joue probablement un rôle limité dans leur génération.
L’urémie semble également réduire de manière importante l’amplitude des ondes de Mayer, ce qui pourrait témoigner d’une altération du fonctionnement du système nerveux autonome cardiovasculaire [117]. Il serait donc intéressant d’étudier les éventuelles variations du MRP chez les patients présentant ce type de pathologie.
En conclusion, les ondes de Traube–Hering et les ondes de Mayer doivent être clairement distinguées. Toutes deux pourraient contribuer au MRP/IRC, sans pour autant correspondre à un seul et même phénomène.
V.2. IRC/MRP et variabilité de la fréquence cardiaque (VFC)
Le cœur ne bat pas avec la régularité d’un métronome : l’intervalle séparant deux battements successifs varie continuellement. Cette variabilité est connue sous le nom de variabilité de la fréquence cardiaque (VFC) ou heart rate variability (HRV) [118]. La VFC est largement reconnue comme un indicateur du fonctionnement du système nerveux autonome (SNA) [119].
De manière générale, une VFC élevée reflète un système nerveux autonome sain et adaptatif, capable d’ajuster rapidement la fréquence cardiaque en réponse aux stimuli internes et externes [120]. À l’inverse, une faible VFC est associée à diverses affections pathologiques, notamment les maladies cardiovasculaires et les neuropathies diabétiques, et témoigne d’une diminution de la capacité d’adaptation du système nerveux autonome [121].
L’analyse de la VFC constitue une méthode non invasive et objective permettant d’évaluer la modulation du SNA et d’apprécier les contributions respectives des activités sympathique et parasympathique. Contrairement à l’idée selon laquelle ces deux branches fonctionneraient exclusivement de manière antagoniste, elles sont aujourd’hui considérées comme agissant de façon coordonnée, parfois en synergie et parfois de manière réciproque [122].
L’analyse spectrale de la VFC est traditionnellement divisée en une bande de basse fréquence (LF ; 0,04–0,15 Hz) et une bande de haute fréquence (HF ; 0,15–0,4 Hz). Toutefois, les limites exactes de ces bandes varient selon les études. La composante HF-VFC, associée à l’arythmie sinusale respiratoire, augmente avec l’activité parasympathique (vagale) [123]. À l’inverse, la composante LF-VFC reflète à la fois des influences sympathiques et parasympathiques et pourrait représenter un mécanisme physiologique indépendant au-delà de sa composante principalement parasympathique [123]. Certains auteurs suggèrent que la bande LF refléterait l’activité sous-harmonique d’un pacemaker central situé dans le tronc cérébral inférieur [123].
Le rapport LF/HF a longtemps été considéré comme un indicateur de l’équilibre sympathovagal. Cette interprétation a cependant été largement remise en question. Dans la mesure où la bande LF ne reflète pas de manière fiable l’activité sympathique, un consensus tend aujourd’hui à considérer que la signification physiologique du rapport LF/HF demeure incertaine, rendant son interprétation particulièrement délicate [124].
Parmi les paramètres les plus fréquemment rapportés figurent la HF-VFC et la LF-VFC. Plus récemment, une bande de fréquence intermédiaire (intermediate frequency band, IM) a été décrite. Cette bande apparaît lors d’états de relaxation hypnoïde et a été mise en évidence dans l’analyse de la perfusion cutanée frontale, des signaux électrocardiographiques et de la respiration [123][125]. Des données préliminaires suggèrent que les oscillations autour de 0,15 Hz prendraient naissance dans le tronc cérébral avant de se propager à la microvascularisation cutanée via les voies parasympathiques [123][125].
Schwerdtfeger et al. [126] ont également proposé que la bande LF puisse en réalité contenir deux rythmes distincts : une composante de plus basse fréquence (0,06–0,1 Hz), principalement associée à la régulation baroréflexe de la pression artérielle via les voies sympathiques, et une composante de fréquence plus élevée (0,1–0,14 Hz), susceptible de refléter des interactions plus complexes entre les activités efférentes sympathique et parasympathique ainsi que les communications cœur–cerveau.
Pelz et al. [9] ont montré que le débit sanguin cutané présentait des profils similaires au MRP/IRC lorsqu’il était mesuré par photopléthysmographie (PPG). Les auteurs suggèrent que le MRP/IRC pourrait correspondre à ce rythme physiologique d’environ 0,15 Hz, également désigné sous le terme de bande intermédiaire (IM). Les oscillations liées à la VFC pourraient ainsi contribuer au phénomène du MRP/IRC [127].
VI. MRP, microcirculation, système lymphatique, matrice extracellulaire et réseau fascial
VI.1. Microcirculation
Environ 10 à 20 % du liquide filtré des capillaires vers l’espace interstitiel retourne à la circulation générale par l’intermédiaire du système lymphatique, tandis que la majorité (80 à 90 %) réintègre les capillaires avant d’être évacuée par le système veineux [128].
Les veinules possèdent un diamètre plus important et des parois musculaires plus fines que les artérioles. Elles sont sous contrôle du système nerveux sympathique, ce qui permet à leurs parois de se contracter ou de se relâcher. Cette propriété joue un rôle majeur dans la capacitance vasculaire et dans la régulation de la perfusion tissulaire. Elle constitue également une base physique à la modulation fréquentielle observée dans les ondes de Traube–Hering et de Mayer [129][130][128].
Sous l’influence des ondes TH et M, et en interaction avec les mécanismes de résistance artérielle, le système de capacitance veineuse subit des contractions lentes et régulières. Ces oscillations favorisent les déplacements de liquide dans l’espace interstitiel, stimulent la circulation lymphatique et facilitent le retour veineux vers le cœur [131][128]. La vasomotricité associée aux oscillations TH et M participe également au maintien de la pression interstitielle négative décrite par l’équilibre de Starling dans les échanges capillaires [128].
Des « ondes de disponibilité en oxygène » oscillantes ont également été décrites. Vern et al. [132] ont rapporté une fréquence moyenne de 9,58 ± 0,117 cycles par minute, tandis que Clark et al. [133] ont observé des fréquences comprises entre 6 et 12 cycles par minute. Ces résultats suggèrent que les variations cycliques du métabolisme oxydatif cortical pourraient constituer un processus oscillatoire local primaire. Ces oscillations métaboliques seraient suivies de modifications hémodynamiques réflexes influençant la perfusion tissulaire locale ainsi que le volume sanguin intracrânien.
Bien que ces processus ne soient pas synchronisés à l’échelle de l’ensemble du cerveau, la proximité des fréquences observées entre les oscillations de l’état rédox du cytochrome oxydase et les ondes THM pourrait permettre leur mise en phase (entrainment). Un tel couplage pourrait assurer un lien entre la régulation métabolique locale et le contrôle hémodynamique central de la perfusion tissulaire [128][134].
VI.2. Systèmes lymphatique et glymphatique, volume cellulaire cérébral et matrice extracellulaire
La pression des fluides tissulaires constitue un déterminant majeur des échanges liquidiens entre les compartiments vasculaire et interstitiel, ainsi qu’entre les espaces interstitiels et les vaisseaux lymphatiques terminaux [135].
Les vaisseaux lymphatiques présentent des contractions rythmiques générées par les cellules musculaires lisses des lymphangions. Ces contractions sont à l’origine d’ondes péristaltiques assurant la progression de la lymphe. Leur fréquence est variable : Mawhinney et Roddie ont rapporté une fréquence moyenne de 2 à 2,5 cycles par minute (extrêmes de 1 à 4 cpm) [136] ; McHale et al. ont observé des fréquences de 4 à 5 cpm dans les lymphatiques mésentériques bovins [137] ; et une fréquence de 4,8 cpm a été décrite dans les vaisseaux lymphatiques poplités du mouton [138]. Plus largement, les vaisseaux lymphatiques peuvent présenter des contractions spontanées comprises entre 1 et 30 cpm [139]. Olszewski [140] a décrit une contractilité lymphatique spontanée variant de 1 à 9 cpm (moyenne d’environ 4 cpm), indépendante du rythme cardiaque, de la respiration ou des mouvements corporels.
Chaque lymphangion est capable de se contracter spontanément de manière autonome grâce à un mécanisme myogène comparable à celui décrit précédemment pour les artérioles [128]. Bien que ces contractions puissent initialement être asynchrones sous l’influence d’une activité pacemaker locale située à proximité des valvules, les lymphangions fonctionnent plus efficacement lorsque leurs contractions deviennent synchronisées. Les vaisseaux lymphatiques présentent d’ailleurs une tendance naturelle à développer facilement une activité coordonnée [141][128]. Une mise en phase (entrainment) induite par les oscillations THM pourrait ainsi optimiser l’efficacité du transport lymphatique.
Les observations expérimentales et cliniques montrent que des contractions lymphatiques rythmiques existent chez de nombreuses espèces, y compris chez l’être humain. Par exemple, le canal thoracique humain se contracte approximativement toutes les 10 à 15 secondes, soit environ quatre pulsations par minute, une fréquence proche de celle décrite pour l’IRC [142]. Sur cette base, Perrin [143] a suggéré que le MRP pourrait correspondre, au moins en partie, à l’expression de contractions lymphatiques rythmiques.
Ferguson [10] considérait cependant peu probable que le système lymphatique soit à l’origine de l’IRC, en raison de l’absence de gros vaisseaux lymphatiques à l’intérieur de la cavité crânienne. Toutefois, les travaux plus récents de Kipnis et de son équipe ont démontré l’existence de vaisseaux lymphatiques associés aux méninges [144]. Cette découverte conduit à s’interroger sur la nécessité de rechercher exclusivement l’origine du MRP au niveau crânien. En effet, certains patients présentent un IRC faible, voire imperceptible au niveau du crâne, alors qu’un rythme clairement perceptible est retrouvé au sacrum ou dans d’autres régions du corps, suggérant une origine plus diffuse.
Le système glymphatique désigne les mécanismes de circulation, de mélange et d’élimination des fluides au sein de l’interstitium cérébral. Ce système comprend trois étapes principales : (1) l’entrée du liquide céphalorachidien (LCR) dans les espaces périvasculaires ; (2) son mélange avec le liquide interstitiel (LI) ; puis (3) l’évacuation de ce mélange hors du tissu cérébral [145].
Les cellules cérébrales, qu’il s’agisse des neurones ou des cellules gliales telles que les astrocytes et les oligodendrocytes, présentent des phénomènes rythmiques de gonflement et de dégonflement liés aux variations intracellulaires de calcium et aux mouvements d’eau qui les accompagnent [16][7][132]. Toutefois, les contractions des oligodendrocytes décrites par Pomerat et al. [146], puis rapportées par Retzlaff et Mitchell [147], se produisent à des fréquences beaucoup plus lentes, de l’ordre de 4 à 18 minutes par cycle.
De façon plus générale, des oscillations intracellulaires du calcium ont été enregistrées à des fréquences comprises entre 0,5 et 12 cycles par minute, aussi bien in vitro qu’in vivo. Ces fréquences sont compatibles avec des rythmes influencés par les oscillations THM et, plus largement, par l’activité du système nerveux autonome [148]. Ces échanges ioniques s’accompagnent également de modifications de la viscosité et des propriétés électriques de la matrice extracellulaire.
La pression de perfusion capillaire, modulée par les ondes THM, pourrait induire des transitions rythmiques entre les états gel et sol de la matrice extracellulaire (MEC), qui se comporte comme un colloïde viscoélastique. Lee [7] décrit la MEC comme un réseau constitué de protéoglycanes (PG), de glycoprotéines (GP) et de glycosaminoglycanes (GAG), formant une structure comparable à un tamis à travers laquelle diffusent les métabolites entre les capillaires et les cellules.
Les protéoglycanes, chargés négativement, fixent l’eau et tendent naturellement à former des structures gélifiées en raison de leur instabilité thermodynamique. Cette organisation confère à la MEC des propriétés comparables à celles d’un cristal liquide ou d’un milieu semi-conducteur. Selon Hamm [77], les modifications cycliques de texture de la MEC pourraient constituer une hypothèse plausible pour expliquer à la fois certaines perceptions palpatoires et certains effets thérapeutiques rapportés en ostéopathie crânienne. Ces propriétés pourraient être modifiées par des tensions fasciales locales, entraînant une augmentation de la densité ou de la rigidité tissulaire.
En résumé, le débit sanguin est influencé non seulement par la compliance et la résistance vasculaires, mais également par les propriétés mécaniques et structurelles des tissus environnants, notamment par la densité et l’organisation locales de la MEC. Sous l’effet de tensions fasciales prolongées, celle-ci peut se densifier par polymérisation des fibres de collagène et accumulation d’agrégats volumineux d’acide hyaluronique [149]. Ces modifications pourraient participer aux mécanismes physiologiques sous-jacents au MRP.
VII. MRP/IRC et synchronisation physiologique (entrainment)
Bien que Ferguson [10] et Perrin [143] aient souligné que les veines cérébrales ne possèdent ni valvules ni couche musculaire lisse significative, et présentent donc peu ou pas de vasomotricité, le rythme perçu par les ostéopathes pourrait provenir de structures vasculaires superficielles et sous-cutanées plutôt que des vaisseaux intracrâniens profonds.
Les ostéopathes décrivent une grande diversité de rythmes appartenant à la plage de fréquences habituellement attribuée au MRP. Il est donc plausible que certains praticiens perçoivent un signal composite résultant de plusieurs oscillations physiologiques superposées, tandis que d’autres n’en détecteraient que certaines composantes. Ces différents rythmes sont probablement intriqués et susceptibles d’interagir les uns avec les autres. Dès lors, la perception palpatoire du MRP/IRC pourrait correspondre à la résultante de plusieurs oscillations physiologiques plutôt qu’à l’expression d’un phénomène unique et isolé.
Cette hypothèse rejoint la théorie proposée par McPartland et Mein [150], selon laquelle le MRP résulterait de la synchronisation — ou entrainment — de multiples rythmes biologiques provenant à la fois du patient et du praticien. Norton [151] a approfondi cette idée à travers son modèle des pressions tissulaires. Selon lui, la perception de l’IRC impliquerait les mécanorécepteurs cutanés à adaptation lente présents chez les deux individus. Les variations de pression tissulaire résulteraient alors de la combinaison des rythmes respiratoires et cardiovasculaires du sujet et de l’examinateur.
Ce modèle pourrait contribuer à expliquer les difficultés observées dans les études de reproductibilité intra- et inter-examinateur. Les différences physiologiques propres à chaque praticien, notamment en termes de fréquence cardiaque ou respiratoire au repos, pourraient influencer le rythme perçu [151].
Tiller et al. [152] ont décrit un phénomène selon lequel différents oscillateurs biologiques tendent à se synchroniser lorsque les activités sympathique et parasympathique sont équilibrées. Dans ces conditions, plusieurs rythmes physiologiques — variabilité de la fréquence cardiaque (HRV), ondes de Traube-Hering-Mayer (THM), respiration, temps de transit du pouls et même certaines activités cérébrales — peuvent s’harmoniser et entrer en résonance. Cette coordination aboutirait à l’émergence d’un rythme dominant que les auteurs désignent sous le terme de fréquence d’entrainment [150].
Chez les sujets en bonne santé, cette fréquence d’entrainment a été mesurée autour de 0,1 Hz, soit environ 6 cycles par minute, une valeur correspondant précisément à la fréquence le plus souvent attribuée au MRP/IRC [152]. Cette convergence suggère que le MRP/IRC pourrait constituer une propriété émergente résultant de la coordination de plusieurs rythmes physiologiques plutôt que d’un mécanisme physiologique unique.
Dans cette perspective, un système nerveux autonome équilibré favoriserait l’émergence d’une oscillation cohérente, stable et sinusoïdale, perceptible sous la forme d’un MRP/IRC ample et régulier. À l’inverse, une dysrégulation autonome pourrait perturber cette coordination et modifier les caractéristiques du MRP, notamment sa fréquence, son amplitude ou sa régularité [150]. Dans les situations de déséquilibre majeur, l’absence de synchronisation entre les différents rythmes physiologiques pourrait même conduire à un MRP/IRC faible, voire imperceptible.
Des études expérimentales complémentaires seront toutefois nécessaires afin de déterminer si cette fréquence d’entrainment peut être mesurée de manière fiable et si elle correspond effectivement au MRP perçu par les praticiens.
VIII. MRP et mouvement embryologique
Un modèle alternatif, généralement désigné sous le terme d’approche biodynamique, a été initialement développé par Rollin Becker dans les années 1930 puis approfondi par James Jealous [150]. Cette approche s’appuie notamment sur les travaux de l’embryologiste Erich Blechschmidt, selon lesquels la fonction, et plus particulièrement le mouvement des fluides, précède et façonne la structure anatomique au cours du développement embryonnaire [154][155].
Blechschmidt observait que la migration et la différenciation cellulaires étaient guidées par des phénomènes dynamiques liés aux mouvements des fluides, qu’il regroupait sous le terme de « biodynamique ». À partir de cette conception, Jealous a formulé l’hypothèse que ces forces organisatrices embryologiques ne disparaissaient pas à la naissance mais continuaient à agir tout au long de la vie, participant aux processus de croissance, d’adaptation et de réparation.
Dans ce cadre théorique, un rythme plus lent que le MRP/IRC a été proposé, souvent désigné comme le rythme à « 2,5 cycles par minute » [150]. La littérature biodynamique introduit également des concepts plus larges et moins précisément définis, tels que le « Souffle de Vie » (Breath of Life) ou la « force thérapeutique », présentés comme des principes organisateurs fondamentaux mais sans fréquence ni origine physiologique clairement établies.
Ces notions dépassant le cadre de cette revue, centrée sur les bases physiologiques du MRP/IRC, elles ne seront pas développées davantage. Elles soulignent néanmoins la possibilité que le MRP/IRC ne constitue pas le rythme biologique le plus fondamental.
Sutherland lui-même évoquait l’existence de rythmes plus profonds, plus subtils et plus difficiles à percevoir que l’IRC [3]. Son élève Rollin Becker [156] décrivait ainsi l’IRC comme un phénomène composé de plusieurs niveaux rythmiques, comprenant notamment une « marée rapide » (fast tide) de 8 à 12 cycles par minute et une « marée lente » (slow tide) d’environ 0,6 cycle par minute [156].
La littérature biodynamique mentionne également une « marée intermédiaire » (mid-tide) de 2 à 3 cycles par minute ainsi qu’une « marée longue » (long tide) d’environ 6 cycles toutes les 9 à 10 minutes, parfois rapprochée du concept de « Souffle de Vie ».
La coexistence de multiples rythmes appartenant à différentes bandes fréquentielles complexifie considérablement l’étude du MRP/IRC. Elle constitue également une explication plausible aux variations observées tant dans les perceptions subjectives que dans les mesures objectives, ainsi qu’aux difficultés de reproductibilité rapportées dans les études intra- et inter-examinateurs.
IX. Implications cliniques et perspectives
Dans cette dernière partie, nous discutons de l’intérêt clinique potentiel de l’IRC/MRP en pratique ostéopathique.
IX.1. Le MRP comme outil ostéopathique
Si les oscillations du MRP reflètent effectivement des variations des propriétés mécaniques du réseau fascial, elles pourraient constituer un outil clinique pertinent pour les thérapeutes manuels cherchant à évaluer et à relâcher les tensions myofasciales. Dans cette perspective, le MRP pourrait servir à la fois d’indicateur diagnostique, en reflétant le niveau de tension tissulaire, et de guide thérapeutique. Ses caractéristiques qualitatives, telles que son amplitude ou sa force, pourraient s’améliorer à mesure que la viscoélasticité et la déformabilité des tissus augmentent.
Afin d’explorer cette hypothèse, il serait intéressant de comparer des mesures objectives à des données palpatoires subjectives. Par exemple, des évaluations réalisées à l’aide d’un Myoton, avec une résolution temporelle élevée, pourraient être corrélées à la palpation du MRP afin de déterminer si des variations cycliques sont observables. Une telle approche permettrait notamment de vérifier si ces oscillations surviennent dans la même gamme de fréquences que le MRP et si elles correspondent à des phénomènes biomécaniques mesurables.
Bien que cette conception soutienne avant tout une approche loco-régionale, la compréhension des mécanismes sous-jacents au MRP renforce également un principe fondamental de l’ostéopathie : l’unité fonctionnelle du corps. Les interventions capables de moduler l’activité du système nerveux autonome (SNA), notamment à travers les interactions cerveau-cœur, pourraient ainsi produire des effets à l’échelle de l’ensemble de l’organisme.
L’ostéopathie manipulative (OMT) est depuis longtemps supposée restaurer certaines fonctions physiologiques en modulant l’activité autonome, favorisant la vasodilatation, diminuant le tonus des muscles lisses et améliorant la circulation [157]. Si le MRP est effectivement lié à la dynamique du SNA, son utilisation comme outil de retour d’information clinique pourrait aider le praticien à apprécier les effets immédiats de son intervention.
La plupart des études rapportent une diminution de l’activité sympathique et une augmentation de la modulation parasympathique après un traitement ostéopathique [124][158], ce qui est cohérent avec une réponse physiologique de relaxation. Il convient toutefois de souligner que ces effets ne sont pas spécifiques à l’ostéopathie. Toute intervention capable d’induire un état de détente — qu’il s’agisse d’un contact manuel non spécifique ou de facteurs contextuels — peut également influencer la régulation autonome [159]. Cette observation souligne l’importance de réaliser des études contrôlées incluant des procédures placebo ou simulées appropriées.
À titre d’exemple, Abenavoli et al. [115] ont évalué les effets de la technique CV4 sur les concentrations de Serum Amyloid A (sAA). Bien qu’ils aient observé une modification physiologique, celle-ci n’était pas significativement différente de celle obtenue avec une procédure simulée. Ce résultat invite à interpréter avec prudence les effets physiologiques attribués à des techniques spécifiques.
La technique CV4 a néanmoins été associée à une diminution du tonus sympathique et à une augmentation de l’activité parasympathique [81][160][19]. Nelson et al. ont montré que le CV4 augmentait l’amplitude des oscillations THM, alors qu’un simple contact crânien n’avait pas cet effet, suggérant l’existence possible d’un effet spécifique au-delà du placebo [161][19]. De même, Sergueef et al. [20] ont montré que certaines techniques crâniennes visant à harmoniser le mouvement crânien global et la jonction cranio-cervicale influençaient les oscillations THM.
Plus récemment, Pelz et al. [9] ont mis en évidence des réponses significatives du système nerveux autonome lors du cranial vault hold (CVH), notamment sur la composante LF de la variabilité cardiaque, sur la bande fréquentielle intermédiaire et sur différents paramètres dérivés de la photopléthysmographie. Les mêmes auteurs ont également observé une augmentation significative du RMSSD après la réalisation d’un CV4 [162], paramètre couramment utilisé comme indicateur de l’activité parasympathique.
D’autres travaux ont utilisé le RMSSD [163], la conductance cutanée [164] ou encore la composante HF de la variabilité cardiaque [165] afin d’objectiver les effets du traitement ostéopathique sur le système nerveux autonome. Fornari et al. [166] ont par ailleurs montré qu’une séance unique d’ostéopathie chez des sujets sains favorisait une récupération plus rapide de la fréquence cardiaque et de l’équilibre sympathovagal après un stress aigu.
Dans leur ensemble, ces résultats suggèrent que l’ostéopathie manipulative pourrait influencer la régulation autonome, notamment en favorisant l’activité parasympathique et en réduisant la prédominance sympathique. Dans ce contexte, le MRP pourrait potentiellement constituer un indicateur clinique de cette modulation autonome.
Traditionnellement, lorsque le corps est considéré comme équilibré, les caractéristiques du MRP — force, amplitude et rythme — sont supposées se situer dans des valeurs jugées normales. Le MRP est alors perçu comme ample, régulier, d’une fréquence proche de 6 cycles par minute chez l’humain, et présentant une expression symétrique entre les phases d’expansion/flexion/rotation externe et de rétraction/extension/rotation interne. Globalement, les tissus semblent osciller entre une phase d’ouverture et une phase de fermeture du corps.
Une symétrie entre les différentes régions corporelles, notamment entre les membres, est également traditionnellement recherchée. Dans la pratique ostéopathique, cette synchronie est souvent interprétée comme le signe d’un équilibre global de l’organisme. À l’inverse, une asymétrie ou une altération des caractéristiques du MRP peut être considérée comme le reflet d’un déséquilibre justifiant une intervention thérapeutique. Lorsque ces paramètres sont jugés satisfaisants, le patient est alors considéré comme rééquilibré et le traitement peut être considéré comme terminé.
Cette notion de synchronie pourrait trouver une explication théorique dans les phénomènes d’entrainment décrits précédemment. Si les différents tissus de l’organisme tendent à s’harmoniser autour d’une fréquence commune, conformément au principe ostéopathique de l’unité du corps, alors une oscillation coordonnée de l’ensemble des tissus pourrait effectivement être attendue.
Cependant, cette notion de synchronisation systémique demeure débattue. Les études palpatoires n’ont pas permis de mettre en évidence de corrélations constantes entre les rythmes perçus à différents sites anatomiques, notamment entre le crâne et le sacrum [170][82][83].
Néanmoins, il demeure possible que l’absence de synchronisation observée dans certaines études soit liée à l’état physiologique des sujets étudiés. Des travaux futurs pourraient évaluer si une meilleure régulation autonome, spontanée ou induite par une intervention ostéopathique, favorise l’émergence d’une synchronisation plus facilement détectable entre différents sites corporels.
Bien qu’il soit possible qu’une telle synchronisation soit plus facilement observable dans certains états physiologiques particulièrement régulés, les données actuelles ne permettent pas de la considérer comme une caractéristique universelle.
IX.2. Idéomotricité
Les phénomènes idéomoteurs correspondent à des micro-mouvements involontaires générés par des processus cognitifs tels que l’attention, l’intention ou l’anticipation, plutôt que par une commande motrice consciente [171].
Appliquée à la palpation ostéopathique, cette notion suggère qu’une partie du MRP perçu pourrait être influencée par des réponses idéomotrices du praticien. Les attentes, le niveau d’attention, l’empathie ou encore la qualité de l’interaction avec le patient peuvent influencer la perception et favoriser ce que la littérature désigne sous le terme d’« expectancy confirmation effects » [172].
Dans leur revue de littérature, Shin et al. [173] reviennent sur plusieurs conceptions historiques de l’idéomotricité, notamment celles développées par William Benjamin Carpenter (1874), Johann Friedrich Herbart (1825) et William James (1890). Bien que ces travaux ne répondent pas aux standards méthodologiques actuels, ils mettent en évidence une idée commune : les représentations mentales et les attentes peuvent influencer directement l’activité motrice, même en l’absence d’intention consciente. Selon Carpenter, le simple fait d’attendre un résultat peut suffire à générer des mouvements involontaires. Herbart suggérait que l’action pouvait être déclenchée par l’anticipation de l’effet sensoriel recherché. Enfin, James considérait que l’imagination d’un mouvement activait déjà la réponse motrice correspondante [173].
Transposée à la pratique ostéopathique, cette théorie conduit à envisager que le fait de rechercher activement un rythme puisse contribuer à le générer ou à en amplifier la perception. L’ostéopathe pourrait ainsi, sans en avoir conscience, participer à la production des micro-mouvements qu’il perçoit ensuite dans les tissus du patient.
Cependant, cette hypothèse ne remet pas nécessairement en cause l’intérêt clinique du phénomène. Les tissus ne peuvent répondre qu’en fonction de leurs propriétés mécaniques intrinsèques. Des tissus souples, peu contraints et présentant de bonnes propriétés viscoélastiques absorberont et transmettront plus facilement ces micro-mouvements, tandis que des tissus plus rigides ou plus densifiés auront tendance à les amortir. Ainsi, même si le mouvement perçu est partiellement influencé par le praticien, la réponse tissulaire reste conditionnée par l’état biomécanique propre du patient.
Dans cette perspective, la palpation du MRP pourrait continuer à fournir des informations pertinentes sur l’état des tissus. L’amplitude, la qualité ou la symétrie de la réponse dépendraient alors davantage des propriétés mécaniques des structures examinées que de l’existence d’un rythme totalement objectif et indépendant de l’observateur. Même lorsqu’elle est influencée par l’action idéomotrice du praticien, la réponse tissulaire demeure limitée par la résilience et la déformabilité des tissus concernés.
Il convient dès lors d’éviter les positions extrêmes. Les sceptiques qui considèrent le MRP/IRC comme une simple hallucination palpatoire ont probablement tort, puisqu’il existe des rythmes physiologiques objectivables et mesurables dans la gamme de fréquences concernée. À l’inverse, les praticiens convaincus que leurs perceptions sont entièrement objectives ont probablement tort également, dans la mesure où les phénomènes idéomoteurs peuvent influencer leur expérience perceptive. Celle-ci pourrait alors résulter d’une superposition complexe de plusieurs rythmes physiologiques présents à différentes profondeurs tissulaires, potentiellement modulés par les rythmes propres du praticien.
De la même manière, la question de la synchronisation mérite une interprétation nuancée. Les praticiens qui considèrent qu’elle ne doit pas être recherchée pourraient négliger une information potentiellement pertinente. Lorsqu’elle est observée, cette synchronisation pourrait en effet témoigner d’une diminution des contraintes tissulaires et d’une meilleure capacité des tissus à répondre aux sollicitations mécaniques induites par l’intention palpatoire du praticien. À l’inverse, considérer cette synchronisation comme un phénomène systématiquement physiologique paraît également excessif. Si le MRP résulte réellement de l’interaction de multiples oscillateurs biologiques, une synchronisation complète, permanente et universelle de l’ensemble des tissus ne saurait être attendue.
IX.3. Cohérence cardiaque, Respiration et MRP
L’Ostéopathie vise à renforcer les capacités d’autorégulation de l’organisme, notamment par une modulation du système nerveux autonome (SNA). Dans cette perspective, l’IRC/MRP pourrait constituer un indicateur clinique reflétant les modifications de l’équilibre autonome.
Pelz et al. [174] ont mis en évidence des réponses rythmiques dans la bande de fréquence intermédiaire (IM) à la suite du Cranial Vault Hold (CVH), une technique couramment utilisée en ostéopathie crânienne. Ces réponses étaient comparables à celles observées lors d’exercices de relaxation autogène, suggérant l’existence de mécanismes physiologiques communs.
Une augmentation de l’amplitude des oscillations perçues du MRP a été proposée comme le reflet d’une meilleure coordination entre les rythmes cardiovasculaires, neurologiques et respiratoires. Plus largement, la synchronisation physiologique (entrainment) est aujourd’hui reconnue comme un mécanisme fondamental des interactions entre le corps et l’esprit. Les approches reposant sur ce phénomène ont montré des effets potentiellement bénéfiques sur les fonctions cognitives, motrices et émotionnelles et font actuellement l’objet d’investigations dans les domaines de la rééducation et de la santé mentale [175].
Certains auteurs ont proposé des modèles plus spéculatifs. Blacklaw-Jones a notamment avancé que des phénomènes piézoélectriques et des champs électromagnétiques endogènes pourraient participer aux effets observés en ostéopathie crânienne [176]. Selon cette hypothèse, les oscillations de l’activité bioélectrique généreraient des vibrations cohérentes au sein des tissus. L’auteur suggère également qu’un praticien entrant dans un état méditatif ou contemplatif pourrait accroître la cohérence de ses propres systèmes physiologiques et, par le contact manuel, influencer l’état physiologique du patient. Ces mécanismes demeurent toutefois théoriques et ne bénéficient actuellement d’aucune validation expérimentale robuste.
Les approches corps-esprit telles que le yoga, la méditation de pleine conscience ou certaines techniques respiratoires ont été associées à une augmentation de la variabilité de la fréquence cardiaque et à une amélioration de l’équilibre autonome [177][36]. Un point commun à nombre de ces pratiques est l’induction d’un état de cohérence cardiaque grâce à des schémas respiratoires contrôlés.
La respiration en cohérence cardiaque consiste généralement à inspirer pendant environ cinq secondes puis à expirer pendant cinq secondes, soit une fréquence respiratoire proche de 0,1 Hz (6 cycles par minute) [179]. Ce rythme respiratoire module directement l’activité autonome : l’inspiration diminue transitoirement l’influence parasympathique et augmente la fréquence cardiaque, tandis que l’expiration renforce l’activité parasympathique et ralentit le rythme cardiaque. Pratiquée régulièrement, cette technique augmente la variabilité de la fréquence cardiaque et améliore la capacité d’adaptation du système nerveux autonome [180][152].
Puisque l’IRC/MRP pourrait résulter de l’intégration de multiples rythmes physiologiques, il est plausible que son amplitude et sa clarté augmentent lors d’états de cohérence autonome. Dans ces conditions, les différents systèmes oscillatoires pourraient se synchroniser davantage, facilitant à la fois la perception palpatoire et les mécanismes de régulation physiologique. Cette harmonisation pourrait également favoriser la circulation des différents fluides de l’organisme, notamment le sang, la lymphe, le liquide céphalo-rachidien et l’air.
On peut dès lors émettre l’hypothèse que la synchronisation des rythmes respiratoires du patient et du praticien au cours de la séance pourrait potentialiser ces effets. Cette hypothèse demeure toutefois spéculative et nécessite d’être validée expérimentalement. Des recherches complémentaires seront nécessaires afin de préciser les liens entre l’IRC/MRP et des processus physiologiques mieux caractérisés, tels que la cohérence cardiaque.
X. Conclusion générale
Cette revue avait pour objectif de réexaminer les mécanismes physiologiques susceptibles de sous-tendre le Mécanisme Respiratoire Primaire (MRP) et l’Impulsion Rythmique Crânienne (IRC).
L’analyse de la littérature montre que le modèle initial proposé par William Garner Sutherland n’est plus pleinement compatible avec les connaissances scientifiques actuelles. Les progrès réalisés en physiologie, en neurosciences et en biomécanique remettent en question les cinq piliers historiques traditionnellement utilisés pour expliquer le MRP.
À l’heure actuelle, aucun mécanisme unique ne permet d’expliquer à lui seul l’ensemble des caractéristiques du MRP/IRC. Les données disponibles soutiennent davantage une interprétation multifactorielle et intégrative. L’hypothèse la plus plausible est que le MRP/IRC émerge de l’interaction de multiples oscillations physiologiques, parmi lesquelles la vasomotricité, l’activité du système nerveux autonome, la variabilité de la fréquence cardiaque, la microcirculation et les dynamiques respiratoires occupent une place centrale.
Dans ce contexte, les variations rythmiques des propriétés viscoélastiques de la matrice extracellulaire, modulées par l’ensemble de ces systèmes interconnectés, pourraient jouer un rôle majeur. Ces fluctuations favoriseraient les échanges métaboliques, le transport des nutriments et l’élimination des déchets, contribuant ainsi au maintien de l’homéostasie tissulaire [181].
Une augmentation de l’amplitude de ces oscillations pourrait refléter une meilleure coordination entre les systèmes cardiovasculaire, neurologique et respiratoire. Cette harmonisation serait susceptible d’améliorer la circulation globale des fluides et de renforcer les capacités d’adaptation de l’organisme.
Plusieurs limites doivent toutefois être soulignées. Les relations entre la perception palpatoire et les phénomènes physiologiques objectivables restent encore imparfaitement comprises. Par ailleurs, l’influence de facteurs liés au praticien, notamment les phénomènes idéomoteurs et les interactions interpersonnelles, ne peut être exclue.
Ces observations plaident en faveur d’un changement de paradigme. Plutôt que de considérer le MRP/IRC comme la conséquence d’un mécanisme unique et strictement mécanique hérité du modèle originel de Sutherland, il semble plus pertinent de l’envisager comme un phénomène émergent résultant de l’interaction dynamique de multiples systèmes biologiques.
Les recherches futures devraient notamment viser à :
- développer des outils de mesure objectifs capables de détecter les oscillations associées au MRP ;
- clarifier les relations entre la palpation et les signaux physiologiques mesurables ;
- évaluer les effets spécifiques des interventions ostéopathiques à l’aide de protocoles expérimentaux rigoureux.
La Figure 2 présente une synthèse schématique des différents mécanismes physiologiques susceptibles de contribuer au phénomène du MRP/IRC.
Conséquence de cette évolution conceptuelle, les auteurs proposent d’abandonner l’expression historiquement chargée de « Mécanisme Respiratoire Primaire » au profit d’une terminologie plus descriptive et factuelle : « Micromouvement Rythmique Palpatoire » (MRP). Cette appellation conserve l’acronyme historique tout en s’affranchissant des présupposés physiologiques du modèle originel.
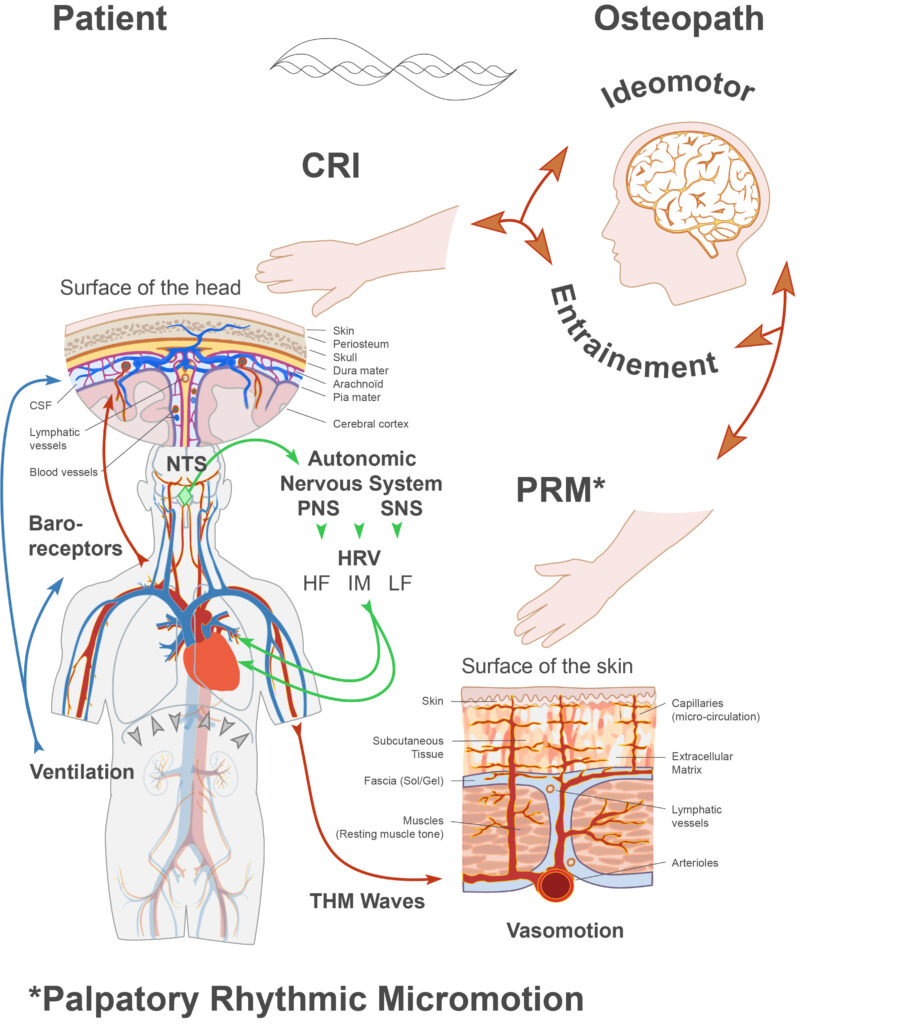
Figure 2. Vue d’ensemble des différents mécanismes sous-jacents au MRP/IRC.
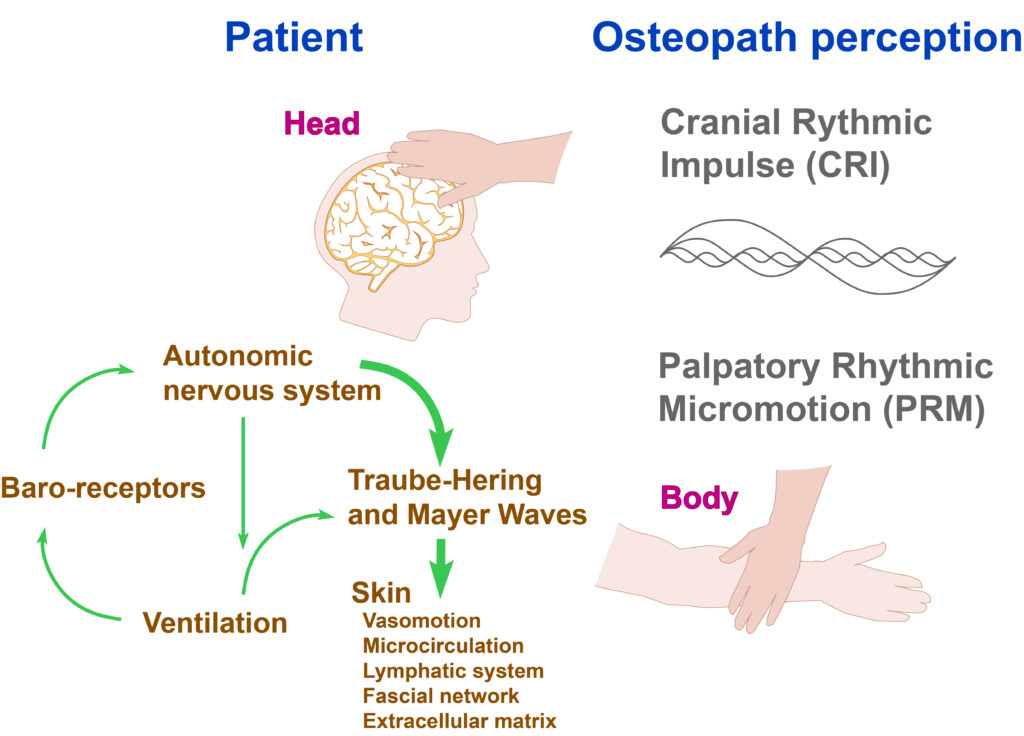
Figure 3. Résumé graphique MRP/IRC
Déclarations
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun intérêt financier ou non financier pertinent à divulguer.
Financement
Aucun financement, subvention ou autre soutien n’a été reçu.
Contribution des auteurs
Tous les auteurs ont contribué à la rédaction de la revue narrative. AG et FM sont à l’origine de l’idée de l’article. LS, AG et FM ont réalisé la recherche bibliographique et l’analyse des données. FM a rédigé la version principale du manuscrit, tandis que LS et AG ont participé à la rédaction et à la révision critique du travail. LS a réalisé une part importante du travail sur les figures. Tous les auteurs ont lu et approuvé la version finale du manuscrit.
Remerciements
Nous remercions chaleureusement Manina Hobson pour sa relecture attentive de l’article en anglais.
Utilisation d’un outil d’intelligence artificielle
Les auteurs ont utilisé ChatGPT (OpenAI) pour les assister dans la révision linguistique de la version anglaise ainsi que dans la traduction française du manuscrit. Cet outil a été utilisé exclusivement afin d’améliorer la lisibilité, la grammaire, le style rédactionnel et la clarté du texte. L’ensemble du contenu scientifique, des analyses, des interprétations, des conclusions et des choix éditoriaux a été élaboré, vérifié et validé par les auteurs, qui assument l’entière responsabilité du manuscrit final.
Titre court :
Mécanismes physiologiques sous-jacents au PRM/CRI en ostéopathie
Références
- a, b, c, d, e, fMériaux F, Stubbe L, Guyon A (2024). « Physiological Mechanisms Underlying the Primary Respiratory Mechanism (PRM) and Cranial Rhythmic Impulse (CRI) in Osteopathy: A Systematic Review. » Healthcare. 12(24):2503. doi:10.3390/healthcare12242503.
- a, b, c, d, e, f, g, h, i, j, k, lMagoun HI (1976). Osteopathy in the Cranial Field. 3rd ed. Sutherland Cranial Teaching Foundation.
- a, b, c, d, e, f, g, h, i, j, k, lSutherland WG (1990). Teachings in the Science of Osteopathy. SCTF.
- a, b, cRasmussen TR, Meulengracht KC (2021). « Direct measurement of the rhythmic motions of the human head identifies a third rhythm. » Journal of Bodywork and Movement Therapies. 26:24–29. doi:10.1016/j.jbmt.2020.08.018.
- a, bWoods JM, Woods RH (1961). « A physical finding related to psychiatric disorders. » The Journal of the American Osteopathic Association. 60:988–993.
- ^Sergueef N, Greer MA, Nelson KE, Glonek T (2011). « The palpated cranial rhythmic impulse (CRI): Its normative rate and examiner experience. » International Journal of Osteopathic Medicine. 14(1):10–16. doi:10.1016/j.ijosm.2010.11.006.
- a, b, cLee RP (2008). « The Living Matrix: A Model for the Primary Respiratory Mechanism. » EXPLORE: The Journal of Science and Healing. 4(6):374–378. doi:10.1016/j.explore.2008.08.003.
- a, bPelz H, Müller G, Keller M, Mathiak K, Mayer J, Borik S, et al. (2023). « Validation of subjective manual palpation using objective physiological recordings of the cranial rhythmic impulse during osteopathic manipulative intervention. » Scientific Reports. 13(1):Article 1. doi:10.1038/s41598-023-33644-8.
- a, b, c, dPelz H, Müller G, Keller M, Mathiak K, Mayer J, Borik S, et al. (2023a). « Validation of subjective manual palpation using objective physiological recordings of the cranial rhythmic impulse during osteopathic manipulative intervention. » Scientific Reports. 13(1):Article 1. doi:10.1038/s41598-023-33644-8.
- a, b, c, d, e, fFerguson A (2003). « A review of the physiology of cranial osteopathy. » Journal of Osteopathic Medicine. 6(2):74–84. doi:10.1016/S1443-8461(03)80017-5.
- a, bGreitz D (1993). « Cerebrospinal fluid circulation and associated intracranial dynamics. A radiologic investigation using MR imaging and radionuclide cisternography. » Acta Radiologica. Supplementum. 386:1–23.
- a, bJordan T (2009). « Swedenborg’s influence on Sutherland’s ‘Primary Respiratory Mechanism’ model in cranial osteopathy. » International Journal of Osteopathic Medicine. 12(3):100–105. doi:10.1016/j.ijosm.2009.03.006.
- a, bMaier SE, Hardy CJ, Jolesz FA (1994). « Brain and cerebrospinal fluid motion: Real-time quantification with M-mode MR imaging. » Radiology. 193(2):477–483. doi:10.1148/radiology.193.2.7972766.
- a, bPoncelet BP, Wedeen VJ, Weisskoff RM, Cohen MS (1992). « Brain parenchyma motion: Measurement with cine echo-planar MR imaging. » Radiology. 185(3):645–651. doi:10.1148/radiology.185.3.1438740.
- a, bUpledger JE (2004). « CranioSacral Therapy. » Seminars in Integrative Medicine. 2(4):159–166. doi:10.1016/j.sigm.2004.12.002.
- a, bFrymann VM (1971). « A study of the rhythmic motions of the living cranium. » The Journal of the American Osteopathic Association. 70(9):928–945.
- a, b, cNelson KE, Sergueef N, Lipinski CM, Chapman AR, Glonek T (2001). « Cranial rhythmic impulse related to the Traube-Hering-Mayer oscillation: Comparing laser-Doppler flowmetry and palpation. » JAOA: Journal of the American Osteopathic Association. 101(3):163–173.
- ^Nelson KE, Sergueef N, Glonek T (2006a). « Recording the Rate of the Cranial Rhythmic Impulse. » JAOA: Journal of the American Osteopathic Association. 106(6):337–341.
- a, b, cNelson KE, Sergueef N, Glonek T (2006b). « The Effect of an Alternative Medical Procedure Upon Low-Frequency Oscillations in Cutaneous Blood Flow Velocity. » Journal of Manipulative and Physiological Therapeutics. 29(8):626–636. doi:10.1016/j.jmpt.2006.08.007.
- a, b, cSergueef N, Nelson KE, Glonek T (2002). « The effect of cranial manipulation on the Traube-Hering-Mayer oscillation as measured by laser-Doppler flowmetry. » Alternative Therapies in Health and Medicine. 8(6):74–76.
- ^Tricco AC, Lillie E, Zarin W, O’Brien KK, Colquhoun H, Levac D, et al. (2018). « PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. » Annals of Internal Medicine. 169(7):467–473. doi:10.7326/M18-0850.
- ^Kellie G (1824). « An Account of the Appearances Observed in the Dissection of Two of Three Individuals Presumed to Have Perished in the Storm of the 3d, and Whose Bodies Were Discovered in the Vicinity of Leith on the Morning of the 4th, November 1821; with Some Reflections on the Pathology of the Brain: Part I. » Transactions. Medico-Chirurgical Society of Edinburgh. 1:84.
- ^Monro A (1783). « Observations on the Structure and Functions of the Nervous System, Illustrated with Tables. » The London Medical Journal. 4(2):113.
- a, bKazimierska A, Manet R, Vallet A, Schmidt E, Czosnyka Z, Czosnyka M, et al. (2023). « Analysis of intracranial pressure pulse waveform in studies on cerebrospinal compliance: A narrative review. » Physiological Measurement. 44(10). doi:10.1088/1361-6579/ad0020.
- ^Brasil S, Solla DJF, Nogueira R de C, Teixeira MJ, Malbouisson LMS, Paiva W da S (2021). « A Novel Noninvasive Technique for Intracranial Pressure Waveform Monitoring in Critical Care. » Journal of Personalized Medicine. 11(12):Article 12. doi:10.3390/jpm11121302.
- ^de Moraes FM, Rocha E, Barros FCD, Freitas FGR, Miranda M, Valiente RA, et al. (2022). « Waveform Morphology as a Surrogate for ICP Monitoring: A Comparison Between an Invasive and a Noninvasive Method. » Neurocritical Care. 37(1):219–227. doi:10.1007/s12028-022-01477-4.
- ^Godoy DA, Brasil S, Iaccarino C, Paiva W, Rubiano AM (2023). « The intracranial compartmental syndrome: A proposed model for acute brain injury monitoring and management. » Critical Care. 27:137. doi:10.1186/s13054-023-04427-4.
- ^Pitlyk PJ, Piantanida TP, Ploeger DW (1985). « Noninvasive intracranial pressure monitoring. » Neurosurgery. 17(4):581–584. doi:10.1227/00006123-198510000-00008.
- ^Kasprowicz M, Lalou DA, Czosnyka M, Garnett M, Czosnyka Z (2016). « Intracranial pressure, its components and cerebrospinal fluid pressure-volume compensation. » Acta Neurologica Scandinavica. 134(3):168–180. doi:10.1111/ane.12541.
- a, b, c, d, eVinje V, Ringstad G, Lindstrøm EK, Valnes LM, Rognes ME, Eide PK, et al. (2019). « Respiratory influence on cerebrospinal fluid flow—A computational study based on long-term intracranial pressure measurements. » Scientific Reports. 9(1):9732. doi:10.1038/s41598-019-46055-5.
- ^Dreha-Kulaczewski S, Joseph AA, Merboldt K-D, Ludwig H-C, Gärtner J, Frahm J (2017). « Identification of the Upward Movement of Human CSF In Vivo and its Relation to the Brain Venous System. » The Journal of Neuroscience. 37(9):2395–2402. doi:10.1523/JNEUROSCI.2754-16.2017.
- a, b, cGutiérrez-Montes C, Coenen W, Vidorreta M, Sincomb S, Martínez-Bazán C, Sánchez AL, et al. (2022). « Effect of Normal Breathing on the Movement of CSF in the Spinal Subarachnoid Space. » AJNR. American Journal of Neuroradiology. 43(9):1369–1374. doi:10.3174/ajnr.A7603.
- ^Mestre H, Tithof J, Du T, Song W, Peng W, Sweeney AM, et al. (2018). « Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. » Nature Communications. 9(1):4878. doi:10.1038/s41467-018-07318-3.
- ^Budgell BS, Bolton PS (2007). « Cerebrospinal Fluid Pressure in the Anesthetized Rat. » Journal of Manipulative and Physiological Therapeutics. 30(5):351–356. doi:10.1016/j.jmpt.2007.04.002.
- ^Conductier G, Brau F, Viola A, Langlet F, Ramkumar N, Dehouck B, et al. (2013). « Melanin-concentrating hormone regulates beat frequency of ependymal cilia and ventricular volume. » Nature Neuroscience. 16(7):845–847. doi:10.1038/nn.3401.
- a, b, c, d, eWhedon JM, Glassey D (2009). « Cerebrospinal fluid stasis and its clinical significance. » Alternative Therapies in Health and Medicine. 15(3):54–60.
- a, bProulx ST (2021). « Cerebrospinal fluid outflow: A review of the historical and contemporary evidence for arachnoid villi, perineural routes, and dural lymphatics. » Cellular and Molecular Life Sciences: CMLS. 78(6):2429–2457. doi:10.1007/s00018-020-03706-5.
- a, bSakka L, Coll G, Chazal J (2011). « Anatomy and physiology of cerebrospinal fluid. » Annales françaises d’Oto-rhino-laryngologie et de Pathologie Cervico-faciale. 128(6):359–366. doi:10.1016/j.aforl.2011.09.004.
- ^Ligocki AP, Vinson AV, Yachnis AT, Dunn WA Jr, Smith DE, Scott EA, et al. (2024). « Cerebrospinal fluid flow extends to peripheral nerves further unifying the nervous system. » Science Advances. 10(36):eadn3259. doi:10.1126/sciadv.adn3259.
- a, bPessa JE (2022). « Ventricular Infusion and Nanoprobes Identify Cerebrospinal Fluid and Glymphatic Circulation in Human Nerves. » Plastic and Reconstructive Surgery. Global Open. 10(2):e4126. doi:10.1097/GOX.0000000000004126.
- a, bDreha-Kulaczewski S, Konopka M, Joseph AA, Kollmeier J, Merboldt K-D, Ludwig H-C, et al. (2018). « Respiration and the watershed of spinal CSF flow in humans. » Scientific Reports. 8(1):Article 1. doi:10.1038/s41598-018-23908-z.
- ^Weller RO (2005). « Microscopic morphology and histology of the human meninges. » Morphologie: Bulletin De l’Association Des Anatomistes. 89(284):22–34. doi:10.1016/s1286-0115(05)83235-7.
- ^Scapinelli R (1990). « Anatomical and radiologic studies on the lumbosacral meningo-vertebral ligaments of humans. » Journal of Spinal Disorders. 3(1):6–15.
- ^Wadhwani S, Loughenbury P, Soames R (2004). « The anterior dural (Hofmann) ligaments. » Spine. 29(6):623–627. doi:10.1097/01.brs.0000115129.59484.24.
- ^Wiltse LL, Fonseca AS, Amster J, Dimartino P, Ravessoud FA (1993). « Relationship of the dura, Hofmann’s ligaments, Batson’s plexus, and a fibrovascular membrane lying on the posterior surface of the vertebral bodies and attaching to the deep layer of the posterior longitudinal ligament. An anatomical, radiologic, and clinical study. » Spine. 18(8):1030–1043. doi:10.1097/00007632-199306150-00013.
- a, b, c, dBordoni B, Walkowski S, Ducoux B, Tobbi F (2020). « The Cranial Bowl in the New Millennium and Sutherland’s Legacy for Osteopathic Medicine: Part 2. » Cureus. 12(9). https://www.cureus.com/articles/40996-the-cranial-bowl-in-the-new-millennium-and-sutherlands-legacy-for-osteopathic-medicine-part-2.
- a, b, c, d, e, f, gAl-Habib A, Albakr A, Al Towim A, Alkubeyyer M, Abu Jamea A, Albadr F, et al. (2018). « In vivo assessment of spinal cord elasticity using shear wave ultrasound in dogs. » Journal of Neurosurgery. Spine. 29(4):461–469. doi:10.3171/2018.2.SPINE171195.
- a, b, c, dRoyo-Salvador MB, Solé-Llenas J, Doménech JM, González-Adrio R (2005). « Results of the section of the filum terminale in 20 patients with syringomyelia, scoliosis and Chiari malformation. » Acta Neurochirurgica. 147(5):515–523; discussion 523. doi:10.1007/s00701-005-0482-y.
- a, b, c, dRoyo-Salvador MB, Fiallos-Rivera MV, Salca HC, Ollé-Fortuny G (2020). « The Filum disease and the Neuro-Cranio-vertebral syndrome: Definition, clinical picture and imaging features. » BMC Neurology. 20:175. doi:10.1186/s12883-020-01743-y.
- a, b, cBordoni B, Zanier E (2015). « Sutherland’s legacy in the new millennium: The osteopathic cranial model and modern osteopathy. » Advances in Mind-Body Medicine. 29(2):15–21.
- ^Clayton EH, Genin GM, Bayly PV (2012). « Transmission, attenuation and reflection of shear waves in the human brain. » Journal of the Royal Society, Interface. 9(76):2899–2910. doi:10.1098/rsif.2012.0325.
- a, bHeisey SR, Adams T (1993). « Role of cranial bone mobility in cranial compliance. » Neurosurgery. 33(5):869–876; discussion 876-877. doi:10.1227/00006123-199311000-00014.
- ^King HH (2007). « The Inherent Rhythmic Motion of the Cranial Bones. » Cranial Letter. 60(1):7–10.
- a, bRetzlaff EW, Michael DK, Roppel RM (1975). « Cranial bone mobility. » The Journal of the American Osteopathic Association. 74(9):869–873.
- a, b, cSeimetz CN, Kemper AR, Duma SM (2012). « An investigation of cranial motion through a review of biomechanically based skull deformation literature. » International Journal of Osteopathic Medicine. 15(4):152–165. doi:10.1016/j.ijosm.2012.05.001.
- ^Rogers JS, Witt PL (1997). « The Controversy of Cranial Bone Motion. » Journal of Orthopaedic & Sports Physical Therapy. 26(2):95–103. doi:10.2519/jospt.1997.26.2.95.
- ^Khandare S, Bhise S, Shinde A (2015). « Age estimation from cranial sutures – a Postmortem study. » International Journal of Healh Care and Biomedical Research. 3–3.
- ^Todd T, Lyon D (1924). « Endocranial Suture Closure: Its Progress and Age Relationship. Part I. » Am. J. Phys. Anthropol. 7:325–384.
- ^Todd T, Lyon D (1925). « Cranial suture closure. Its progress and age relationship. Part III.—Endocranial closure in adult males of Negro stock. » Am. J. Phys. Anthropol. 8:47–71.
- a, bBassed RB, Briggs C, Drummer OH (2010). « Analysis of time of closure of the spheno-occipital synchondrosis using computed tomography. » Forensic Science International. 200(1–3):161–164. doi:10.1016/j.forsciint.2010.04.009.
- a, bStarkey R (2015). « Arguments for and against movement at the spheno-occipital synostosis: Furthering the debate. » International Journal of Osteopathic Medicine. 18(2):102–115. doi:10.1016/j.ijosm.2014.12.001.
- ^Madeline LA, Elster AD (1995). « Suture closure in the human chondrocranium: CT assessment. » Radiology. 196(3):747–756. doi:10.1148/radiology.196.3.7644639.
- ^Shirley NR, Jantz RL (2011). « Spheno-occipital synchondrosis fusion in modern Americans. » Journal of Forensic Sciences. 56(3):580–585. doi:10.1111/j.1556-4029.2011.01705.x.
- ^Downey PA, Barbano T, Kapur-Wadhwa R, Sciote JJ, Siegel MI, Mooney MP (2006). « Craniosacral Therapy: The Effects of Cranial Manipulation on Intracranial Pressure and Cranial Bone Movement. » Journal of Orthopaedic & Sports Physical Therapy. 36(11):845–853. doi:10.2519/jospt.2006.36.11.845.
- ^Dechant JJ, Mooney MP, Cooper GM, Smith TD, Burrows AM, Losken HW, et al. (1999). « Positional changes of the frontoparietal ossification centers in perinatal craniosynostotic rabbits. » Journal of Craniofacial Genetics and Developmental Biology. 19(2):64–74.
- ^Greenman PE, McPartland JM (1995). « Cranial findings and iatrogenesis from craniosacral manipulation in patients with traumatic brain syndrome. » JAOA: Journal of the American Osteopathic Association. 95(3):182–182.
- ^Oleski SL, Smith GH, Crow WT (2002). « Radiographic evidence of cranial bone mobility. » Cranio: The Journal of Craniomandibular Practice. 20(1):34–38. doi:10.1080/08869634.2002.11746188.
- ^Cook A (2005). « The mechanics of cranial motion—The sphenobasilar synchondrosis (SBS) revisited. » Journal of Bodywork and Movement Therapies. 9(3):177–188. doi:10.1016/j.jbmt.2004.12.002.
- ^Crow WT, King HH, Patterson RM, Giuliano V (2009). « Assessment of calvarial structure motion by MRI. » Osteopathic Medicine and Primary Care. 3(1):8. doi:10.1186/1750-4732-3-8.
- a, b, cGabutti M, Draper-Rodi J (2014). « Osteopathic decapitation: Why do we consider the head differently from the rest of the body? New perspectives for an evidence informed osteopathic approach to the head. » International Journal of Osteopathic Medicine. doi:10.1016/j.ijosm.2014.02.001.
- ^Herring SW, Teng S (2000). « Strain in the braincase and its sutures during function. » American Journal of Physical Anthropology. 112(4):575–593. doi:10.1002/1096-8644(200008)112:4<575::AID-AJPA10>3.0.CO;2-0.
- ^Jaslow CR (1990). « Mechanical properties of cranial sutures. » Journal of Biomechanics. 23(4):313–321. doi:10.1016/0021-9290(90)90059-C.
- ^Maloul A, Fialkov J, Whyne CM (2013). « Characterization of the bending strength of craniofacial sutures. » Journal of Biomechanics. 46(5):912–917. doi:10.1016/j.jbiomech.2012.12.016.
- ^Wang J-W, Yu K, Li M, Wu J, Wang J, Wan C-W, et al. (2022). « Application of nanoindentation technology in testing the mechanical properties of skull materials. » Scientific Reports. 12(1):8717. doi:10.1038/s41598-022-11216-6.
- ^Adams T, Heisey RS, Smith MC, Briner BJ (1992). « Parietal bone mobility in the anesthetized cat. » The Journal of the American Osteopathic Association. 92(5):599–600, 603–610, 615–622.
- a, b, cMoskalenko YE, Frymann V, Kravchenko T, Weinstein GB (2003). « Physiological Background of the Cranial Rhythmic Impulse and The Primary Respiratory Mechanism. » AAO Journal. 13(2):21–33.
- a, b, c, d, e, fHamm D (2011). « A hypothesis to explain the palpatory experience and therapeutic claims in the practice of osteopathy in the cranial field. » International Journal of Osteopathic Medicine. 14(4):149–165. doi:10.1016/j.ijosm.2011.07.003.
- ^Troup JDG (1986). « Biomechanics of the lumbar spinal canal. » Clinical Biomechanics. 1(1):31–43. doi:10.1016/0268-0033(86)90036-7.
- ^Dai LY, Xu YK, Zhang WM, Zhou ZH (1989). « The effect of flexion-extension motion of the lumbar spine on the capacity of the spinal canal. An experimental study. » Spine. 14(5):523–525.
- ^Cella M, Acella E, Aquino A, Pisa V (2022). « Cranial osteopathic techniques and electroencephalogram (EEG) alpha power: A controlled crossover trial. » Journal of Osteopathic Medicine. 122(8):401–409. doi:10.1515/jom-2021-0257.
- a, bMiana L, Hugo do Vale Bastos V, Machado S, Arias-Carrión O, Nardi AE, Almeida L, et al. (2013). « Changes in alpha band activity associated with application of the compression of fourth ventricular (CV-4) osteopathic procedure: A qEEG pilot study. » Journal of Bodywork and Movement Therapies. 17(3):291–296. doi:10.1016/j.jbmt.2012.10.002.
- a, bMoran RW, Gibbons P (2001). « Intraexaminer and interexaminer reliability for palpation of the cranial rhythmic impulse at the head and sacrum. » Journal of Manipulative and Physiological Therapeutics. 24(3):183–190. doi:10.1016/S0161-4754(01)62711-8.
- a, bRogers JS, Witt PL, Gross MT, Hacke JD, Genova PA (1998). « Simultaneous palpation of the craniosacral rate at the head and feet: Intrarater and interrater reliability and rate comparisons. » Physical Therapy. 78(11):1175–1185. doi:10.1093/ptj/78.11.1175.
- ^Jones TW (1853). « Discovery That the Veins of the Bat’s Wing (Which Are Furnished with Valves) Are Endowed with Rhythmical Contractility, and That the Onward Flow of Blood Is Accelerated by Such Contraction. » Edinburgh Medical and Surgical Journal. 79(195):367–373.
- ^Intaglietta M (1991). « Arteriolar vasomotion: Implications for tissue ischemia. » Blood Vessels. 28 Suppl 1:1–7. doi:10.1159/000158912.
- a, bMileva N, Velikova T, Velikov T, Vassilev D (2024). « Aortic Elasticity and Cardiovascular Risk Stratification: A Narrative Review on the Current Understanding. » Journal of Vascular Diseases. 3(1):88–101. doi:10.3390/jvd3010008.
- a, bFredriksson I, Larsson M, Strömberg T, Iredahl F (2021). « Vasomotion analysis of speed resolved perfusion, oxygen saturation, red blood cell tissue fraction, and vessel diameter: Novel microvascular perspectives. » Skin Research and Technology. 28(1):142–152. doi:10.1111/srt.13106.
- a, bStefanovska A, Bracic M, Kvernmo HD (1999). « Wavelet analysis of oscillations in the peripheral blood circulation measured by laser Doppler technique. » IEEE Transactions on Biomedical Engineering. 46(10):1230–1239. doi:10.1109/10.790500.
- ^Chaudhry R, Miao JH, Rehman A (2025). « Physiology, Cardiovascular. » In StatPearls. StatPearls Publishing. http://www.ncbi.nlm.nih.gov/books/NBK493197/.
- a, b, cPappano AJ, Gil Wier W (2013). « 9—The Peripheral Circulation and its Control. » In Pappano AJ, Gil Wier W (Eds.), Cardiovascular Physiology. 10th ed. Elsevier. pp. 171–194. doi:10.1016/B978-0-323-08697-4.00009-5.
- a, b, c, d, e, f, gNilsson H, Aalkjaer C (2003). « Vasomotion: Mechanisms and physiological importance. » Molecular Interventions. 3(2):79–89. doi:10.1124/mi.3.2.79.
- ^Peng H, Matchkov V, Ivarsen A, Aalkjaer C, Nilsson H (2001). « Hypothesis for the initiation of vasomotion. » Circulation Research. 88(8):810–815. doi:10.1161/hh0801.089603.
- a, bColantuoni A, Bertuglia S, Coppini G, Donato L (1990). « Superposition of arteriolar vasomotion waves and regulation of blood flow in skeletal muscle microcirculation. » Advances in Experimental Medicine and Biology. 277:549–558. doi:10.1007/978-1-4684-8181-5_62.
- a, bAon MA, Cortassa S, O’Rourke B (2008). « Mitochondrial Oscillations in Physiology and Pathophysiology. » Advances in Experimental Medicine and Biology. 641:98–117. doi:10.1007/978-0-387-09794-7_8.
- ^Cervera J, Meseguer S, Mafe S (2018). « Intercellular Connectivity and Multicellular Bioelectric Oscillations in Nonexcitable Cells: A Biophysical Model. » ACS Omega. 3(10):13567–13575. doi:10.1021/acsomega.8b01514.
- ^Figueroa XF, Duling BR (2009). « Gap Junctions in the Control of Vascular Function. » Antioxidants & Redox Signaling. 11(2):251–266. doi:10.1089/ars.2008.2117.
- ^Aalkjaer C, Nilsson H (2005). « Vasomotion: Cellular background for the oscillator and for the synchronization of smooth muscle cells. » British Journal of Pharmacology. 144(5):605–616. doi:10.1038/sj.bjp.0706084.
- a, bVillermain-Lecolier G, Billaudel P, Perrin E, Scherpereel B, Pire J-C (1994). « Detection of periodic micro movements using a non destructive method. » Rbm -Paris-. https://hal.science/hal-02334851.
- ^Billaudel P, Lecolier GV, Pire J, Laval Y (1991). « Detection of periodic micro movements of the head using a flat sensors system. » [1991 Proceedings] 6th Mediterranean Electrotechnical Conference. 752–755 vol.1. doi:10.1109/MELCON.1991.161947.
- 100.a, bStill AT (1897). Autobiography of Andrew T. Still, with a history of the discovery and development of the science of osteopathy, together with an account of the founding of the American school of osteopathy. Kirksville, Mo.: The author. http://archive.org/details/autobiographyand00stiliala.
- 101.^Barnett WH, Latash EM, Capps RA, Dick TE, Wehrwein EA, Molkov YI (2020). « Traube-Hering waves are formed by interaction of respiratory sinus arrhythmia and pulse pressure modulation in healthy men. » Journal of Applied Physiology. 129(5):1193–1202. doi:10.1152/japplphys.00452.2020.
- 102.a, b, c, dJulien C (2006). « The enigma of Mayer waves: Facts and models. » Cardiovascular Research. 70(1):12–21. doi:10.1016/j.cardiores.2005.11.008.
- 103.^Julien C (2008). « Baroreflex Control of Sympathetic Nerve Activity and Blood Pressure Variability. » Clinical and Experimental Pharmacology and Physiology. 35(4):512–515. doi:10.1111/j.1440-1681.2008.04907.x.
- 104.^deBoer R, Karemaker J, J S (1987). « Hemodynamic fluctuations and baroreflex sensitivity in humans: A beat-to-beat model. » The American Journal of Physiology. 253(3 Pt 2). doi:10.1152/ajpheart.1987.253.3.H680.
- 105.^La Fountaine M, Hohn A, Leahy C, Testa AJ, Weir JP (2022). « Use of Mayer wave activity to demonstrate aberrant cardiovascular autonomic control following sports concussion injury. » Annals of the New York Academy of Sciences. 1507(1). doi:10.1111/nyas.14683.
- 106.^Finucane C, Boyle G, Fan CW, Hade D, Byrne L, Kenny RA (2010). « Mayer wave activity in vasodepressor carotid sinus hypersensitivity. » Europace. 12(2):247–253. doi:10.1093/europace/eup385.
- 107.a, bJulien C (2019). « Physiology of low-frequency heart rate oscillations. » J. Physiol. 597(10):2595–2598. doi:10.1113/JP277500.
- 108.a, bGrasso R, Schena F, Gulli G, Cevese A (1997). « Does low-frequency variability of heart period reflect a specific parasympathetic mechanism? » Journal of the Autonomic Nervous System. 63(1):30–38. doi:10.1016/S0165-1838(96)00128-2.
- 109.^Fernandez D, Lecine A (1990). « Recording of the Traube-Hering wave and simultaneous cranial palpation. » Kinésithérapie scientifique. 292:33–40.
- ^Guinn K, Ali H, Seffinger M (2006). « Validation of Transcutaneous Laser Doppler Flowmeter in Measuring Autonomic Tone. » Journal of Investigative Medicine. 54(1):S172–S172. doi:10.2310/6650.2005.X0004.541.
- a, bChrist F, Raithel P, Gartside IB, Gamble J, Peter K, Messmer K (1995). « Investigating the origin of cyclic changes in limb volume using mercury-in-silastic strain gauge plethysmography in man. » The Journal of Physiology. 487(1):259–272. doi:10.1113/jphysiol.1995.sp020877.
- ^Baskin RJ, Paolini PJ (1966). « Muscle volume changes. » The Journal of General Physiology. 49(3):387–404. doi:10.1085/jgp.49.3.387.
- ^Scarr G (2016). « Fascial hierarchies and the relevance of crossed-helical arrangements of collagen to changes in shape; part II: The proposed effect of blood pressure (Traube-Hering-Mayer) waves on the fascia. » Journal of Bodywork and Movement Therapies. 20(3):629–638. doi:10.1016/j.jbmt.2015.10.008.
- ^Abenavoli A, Pisa S, Maggiani A (2020b). « A Pilot Study of Jugular Compression (Queckenstedt maneuver) for Cranial Movement Perception. » The Journal of the American Osteopathic Association. 120(10):647–654. doi:10.7556/jaoa.2020.119.
- a, b, cAbenavoli A, Badi F, Barbieri M, Bianchi M, Biglione G, Dealessi C, et al. (2020). « Cranial osteopathic treatment and stress-related effects on autonomic nervous system measured by salivary markers: A pilot study. » J Bodyw Mov Ther. 24(4):215–221. doi:10.1016/j.jbmt.2020.07.017.
- ^Taylor JA, Carr DL, Myers CW, Eckberg DL (1998). « Mechanisms underlying very-low-frequency RR-interval oscillations in humans. » Circulation. 98(6):547–555. doi:10.1161/01.cir.98.6.547.
- ^Cloarec-Blanchard L, Girard A, Houhou S, Grünfeld JP, Elghozi JL (1992). « Spectral analysis of short-term blood pressure and heart rate variability in uremic patients. » Kidney International. Supplement. 37:S14-18.
- ^Rajendra Acharya U, Paul Joseph K, Kannathal N, Lim CM, Suri JS (2006). « Heart rate variability: A review. » Medical & Biological Engineering & Computing. 44(12):1031–1051. doi:10.1007/s11517-006-0119-0.
- ^Stein PK, Bosner MS, Kleiger RE, Conger BM (1994). « Heart rate variability: A measure of cardiac autonomic tone. » American Heart Journal. 127(5):1376–1381. doi:10.1016/0002-8703(94)90059-0.
- 120.^Tiwari R, Kumar R, Malik S, Raj T, Kumar P (2021). « Analysis of Heart Rate Variability and Implication of Different Factors on Heart Rate Variability. » Current Cardiology Reviews. 17(5):e160721189770. doi:10.2174/1573403X16999201231203854.
- 121.^Pumprla J, Howorka K, Groves D, Chester M, Nolan J (2002). « Functional assessment of heart rate variability: Physiological basis and practical applications. » International Journal of Cardiology. 84(1):1–14. doi:10.1016/S0167-5273(02)00057-8.
- 122.^Ondicova K, Mravec B (2010). « Multilevel interactions between the sympathetic and parasympathetic nervous systems: A minireview. » Endocrine Regulations. 44(2):69–75. doi:10.4149/endo_2010_02_69.
- 123.a, b, c, d, eKeller M, Pelz H, Perlitz V, Zweerings J, Röcher E, Baqapuri HI, et al. (2020). « Neural correlates of fluctuations in the intermediate band for heart rate and respiration are related to interoceptive perception. » Psychophysiology. 57(9):e13594. doi:10.1111/psyp.13594.
- 124.a, bCarnevali L, Lombardi L, Fornari M, Sgoifo A (2020). « Exploring the Effects of Osteopathic Manipulative Treatment on Autonomic Function Through the Lens of Heart Rate Variability. » Frontiers in Neuroscience. 14:579365. doi:10.3389/fnins.2020.579365.
- 125.a, bPerlitz V, Lambertz M, Cotuk B, Grebe R, Vandenhouten R, Flatten G, et al. (2004). « Cardiovascular rhythms in the 0.15-Hz band: Common origin of identical phenomena in man and dog in the reticular formation of the brain stem? » Pflügers Archiv. 448(6):579–591. doi:10.1007/s00424-004-1291-4.
- 126.^Schwerdtfeger AR, Schwarz G, Pfurtscheller K, Thayer JF, Jarczok MN, Pfurtscheller G (2020). « Heart rate variability (HRV): From brain death to resonance breathing at 6 breaths per minute. » Clinical Neurophysiology: Official Journal of the International Federation of Clinical Neurophysiology. 131(3):676–693. doi:10.1016/j.clinph.2019.11.013.
- 127.^Keller M, Perlitz V, Pelz H, Borik S, Repik I, Geilgens A, et al. (2025). « Specificity of cranial cutaneous manipulations in modulating autonomic nervous system responses and physiological oscillations: A controlled study. » PloS One. 20(2):e0317300. doi:10.1371/journal.pone.0317300.
- 128.a, b, c, d, e, f, gNelson KE (2002). « The primary respiratory mechanism. » 12(4):25–34.
- 129.^Gretchin VB (1969). « Some data on oxygen dynamics in subcortical structures of the human brain. » Electroencephalography and Clinical Neurophysiology. 26(5):546–547.
- 130.^Kobayashi M, Musha T (1982). « 1/f fluctuation of heartbeat period. » IEEE Transactions on Bio-Medical Engineering. 29(6):456–457. doi:10.1109/TBME.1982.324972.
- 131.^Intaglietta M (1981). « Vasomotor activity, time-dependent fluid exchange and tissue pressure. » Microvascular Research. 21(2):153–164. doi:10.1016/0026-2862(81)90028-5.
- 132.a, bVern BA, Leheta BJ, Juel VC, LaGuardia J, Graupe P, Schuette WH (1998). « Slow oscillations of Cytochrome Oxidase Redox State and Blood Volume in Unanesthetized Cat and Rabbit Cortex. » Advances in Experimental Medicine and Biology. 454:561–570. doi:10.1007/978-1-4615-4863-8_67.
- 133.^Clark LC, Misrahy G, Fox RP (1958). « Chronically implanted polarographic electrodes. » J Appl Physiol. 13(1):85–91. doi:10.1152/jappl.1958.13.1.85.
- 134.^Steinmeier R, Bauhuf C, Hübner U, Bauer RD, Fahlbusch R, Laumer R, Bondar I (1996). « Slow rhythmic oscillations of blood pressure, intracranial pressure, microcirculation, and cerebral oxygenation. Dynamic interrelation and time course in humans. » Stroke. 27(12):2236–2243. doi:10.1161/01.str.27.12.2236.
- 135.^Breslin JW, Yang Y, Scallan JP, Sweat RS, Adderley SP, Murfee WL (2018). « Lymphatic Vessel Network Structure and Physiology. » Comprehensive Physiology. 9(1):207–299. doi:10.1002/cphy.c180015.
- 136.a, bMawhinney HJ, Roddie IC (1973). « Spontaneous activity in isolated bovine mesenteric lymphatics. » The Journal of Physiology. 229(2):339–348. doi:10.1113/jphysiol.1973.sp010141.
- 137.^McHale NG, Roddie IC (1976). « The effect of transmural pressure on pumping activity in isolated bovine lymphatic vessels. » The Journal of Physiology. 261(2):255–269.
- 138.^McGeown JG, McHale NG, Thornbury KD (1987). « The role of external compression and movement in lymph propulsion in the sheep hind limb. » The Journal of Physiology. 387:83–93. doi:10.1113/jphysiol.1987.sp016564.
- 139.^Hall JG, Morris B, Woolley G (1965). « Intrinsic rhythmic propulsion of lymph in the unanaesthetized sheep. » The Journal of Physiology. 180(2):336–349. doi:10.1113/jphysiol.1965.sp007706.
- 140.^Olszewski WL (2002). « Contractility patterns of normal and pathologically changed human lymphatics. » Annals of the New York Academy of Sciences. 979:52–63. doi:10.1111/j.1749-6632.2002.tb04867.x.
- 141.^Degenhardt BF, Kuchera ML (1996). « Update on osteopathic medical concepts and the lymphatic system. » The Journal of the American Osteopathic Association. 96(2):97–100. doi:10.7556/jaoa.1996.96.2.97.
- 142.^Kinmonth JB, Taylor GW, Tracy GD, Marsh JD (1957). « Primary lymphoedema; clinical and lymphangiographic studies of a series of 107 patients in which the lower limbs were affected. » The British Journal of Surgery. 45(189):1–9. doi:10.1002/bjs.18004518902.
- 143.a, bPerrin RN (2007). « Lymphatic Drainage of the Neuraxis in Chronic Fatigue Syndrome: A Hypothetical Model for the Cranial Rhythmic Impulse. » JAOA: Journal of the American Osteopathic Association. 107(6):218–224.
- 144.^Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, et al. (2015). « Structural and functional features of central nervous system lymphatic vessels. » Nature. 523(7560):337–341. doi:10.1038/nature14432.
- 145.^Bohr T, Hjorth PG, Holst SC, Hrabětová S, Kiviniemi V, Lilius T, et al. (2022). « The glymphatic system: Current understanding and modeling. » iScience. 25(9):104987. doi:10.1016/j.isci.2022.104987.
- 146.a, bPomerat CM (1959). « Rhythmic contraction of Schwann cells. » Science (New York, N.Y.). 130(3391):1759–1760. doi:10.1126/science.130.3391.1759-a.
- 147.a, bRetzlaff EW (1987). « Anatomy and Physiology of Craniosacral Mechanisms. » In Retzlaff EW, Mitchell FL (Eds.), The Cranium and Its Sutures: Anatomy, Physiology, Clinical Applications and Annotated Bibliography of Research in the Cranial Field. Springer. pp. 5–12. doi:10.1007/978-3-642-71848-9_2.
- 148.^Strumwasser F, Vogel JM (1992). « Cellular oscillators and biological timing: The role of proteins and Ca2+. » Progress in Brain Research. 92:309–320. doi:10.1016/s0079-6123(08)61185-1.
- 149.^Stecco A, Cowman M, Pirri N, Raghavan P, Pirri C (2022). « Densification: Hyaluronan Aggregation in Different Human Organs. » Bioengineering (Basel, Switzerland). 9(4):159. doi:10.3390/bioengineering9040159.
- 150.a, b, c, d, eMcPartland JM, Mein EA (1997). « Entrainment and the cranial rhythmic impulse. » Alternative Therapies in Health and Medicine. 3(1):40–45.
- 151.a, bNorton JM (1991). « A tissue pressure model for palpatory perception of the cranial rhythmic impulse. » JAOA: Journal of the American Osteopathic Association. 91(10):975–975.
- 152.a, b, c, dTiller WA, McCraty R, Atkinson M (1996). « Cardiac coherence: A new, noninvasive measure of autonomic nervous system order. » Alternative Therapies in Health and Medicine. 2(1):52–65.
- 153.^Greenman PE, Mein EA, Andary M (1996). « Craniosacral Manipulation. » Physical Medicine and Rehabilitation Clinics of North America. 7(4):877–896. doi:10.1016/S1047-9651(18)30371-1.
- 154.^Blechschmidt E (2004). The Ontogenetic Basis of Human Anatomy: A Biodynamic Approach to Development from Conception to Birth. North Atlantic Books.
- 155.^Blechschmidt E, Gasser R (2012). Biokinetics and Biodynamics of Human Differentiation: Principles and Applications. 2nd ed. North Atlantic Books.
- 156.a, bBecker RE (1997). Life in Motion. Portland, OR: Rudra Press.
- 157.^Rechberger V, Biberschick M, Porthun J (2019). « Effectiveness of an osteopathic treatment on the autonomic nervous system: A systematic review of the literature. » European Journal of Medical Research. 24(1):36. doi:10.1186/s40001-019-0394-5.
- 158.^Henley CE, Ivins D, Mills M, Wen FK, Benjamin BA (2008). « Osteopathic manipulative treatment and its relationship to autonomic nervous system activity as demonstrated by heart rate variability: A repeated measures study. » Osteopathic Medicine and Primary Care. 2:7. doi:10.1186/1750-4732-2-7.
- 159.^Tarsha MS, Park S, Tortora S (2019). « Body-Centered Interventions for Psychopathological Conditions: A Review. » Frontiers in Psychology. 10:2907. doi:10.3389/fpsyg.2019.02907.
- 160.^Milnes K, Moran RW (2007). « Physiological effects of a CV4 cranial osteopathic technique on autonomic nervous system function: A preliminary investigation. » International Journal of Osteopathic Medicine. 10(1):8–17. doi:10.1016/j.ijosm.2007.01.003.
- 161.^Nelson K, Sergueef N, Glonek T (2004). « Cranial manipulation induces sequential changes in blood flow velocity on demand. » AAO Journal. 14(3):15–17.
- 162.^Keller M, Perlitz V, Pelz H, Borik S, Repik I, Geilgens A, et al. (2024). On the specificity of manual triggers of autonomic nervous system responses using osteopathic cranial manipulative medicine. bioRxiv. https://doi.org/10.1101/2024.09.11.612567.
- 163.^Besson C, Mur T, Benaim C, Schmitt L, Gremeaux V (2023). « Short-term effects on heart rate variability of occipito-mastoid suture normalization in healthy subjects. » Frontiers in Neuroscience. 17:1271461. doi:10.3389/fnins.2023.1271461.
- 164.^Arienti C, Farinola F, Ratti S, Daccò S, Fasulo L (2020). « Variations of HRV and skin conductance reveal the influence of CV4 and Rib Raising techniques on autonomic balance: A randomized controlled clinical trial. » Journal of Bodywork and Movement Therapies. 24(4):395–401. doi:10.1016/j.jbmt.2020.07.002.
- 165.^Ruffini N, D’Alessandro G, Mariani N, Pollastrelli A, Cardinali L, Cerritelli F (2015). « Variations of high frequency parameter of heart rate variability following osteopathic manipulative treatment in healthy subjects compared to control group and sham therapy: Randomized controlled trial. » Frontiers in Neuroscience. 9:272. doi:10.3389/fnins.2015.00272.
- 166.^Fornari M, Carnevali L, Sgoifo A (2017). « Single Osteopathic Manipulative Therapy Session Dampens Acute Autonomic and Neuroendocrine Responses to Mental Stress in Healthy Male Participants. » The Journal of the American Osteopathic Association. 117(9):559–567. doi:10.7556/jaoa.2017.110.
- 167.^Bayo-Tallón V, Esquirol-Caussa J, Pàmias-Massana M, Planells-Keller K, Palao-Vidal DJ (2019). « Effects of manual cranial therapy on heart rate variability in children without associated disorders: Translation to clinical practice. » Complementary Therapies in Clinical Practice. 36:125–141. doi:10.1016/j.ctcp.2019.06.008.
- 168.^Byrne EA, Porges SW (1992). « Frequency-specific amplification of heart rate rhythms using oscillatory tilt. » Psychophysiology. 29(1):120–126. doi:10.1111/j.1469-8986.1992.tb02022.x.
- 169.^Tamburella F, Piras F, Piras F, Spanò B, Tramontano M, Gili T (2019). « Cerebral Perfusion Changes After Osteopathic Manipulative Treatment: A Randomized Manual Placebo-Controlled Trial. » Frontiers in Physiology. 10:403. doi:10.3389/fphys.2019.00403.
- 170.^Burch GE, Cohn AE, Neumann C (1942). « A study by quantitative methods of the spontaneous variations in volume of the finger tip, toe tip, and postero-superior portion of the pinna of resting normal white adults. » American Journal of Physiology-Legacy Content. 136(3):433–447. doi:10.1152/ajplegacy.1942.136.3.433.
- 171.^Dorko BL (2003). « The analgesia of movement: Ideomotor activity and manual care. » Journal of Osteopathic Medicine. 6(2):93–95. doi:10.1016/S1443-8461(03)80022-9.
- 172.^Spitz HH (1997). Nonconscious movements: From mystical messages to facilitated communication. Lawrence Erlbaum Associates, Inc. pp. xii, 202.
- 173.a, b, cShin YK, Proctor RW, Capaldi EJ (2010). « A review of contemporary ideomotor theory. » Psychological Bulletin. 136(6):943–974. doi:10.1037/a0020541.
- 174.^Pelz H, Keller M, Müller G, Mathiak K, Mayer J, Perlitz V (2022). « Immediate autonomic nervous system activity in skin microcirculation during osteopathic cranial vault hold intervention. » Research Square. doi:10.21203/rs.3.rs-2409912/v1.
- 175.^Barbaresi M, Nardo D, Fagioli S (2025). « Physiological Entrainment: A Key Mind–Body Mechanism for Cognitive, Motor and Affective Functioning, and Well-Being. » Brain Sciences. 15(1):3. doi:10.3390/brainsci15010003.
- 176.a, bBlacklaw-Jones RD (2009). « Comments on cranio-sacral method and efficacy. » International Journal of Osteopathic Medicine. 12(2):72–74. doi:10.1016/j.ijosm.2008.10.001.
- 177.^Mehling WE, Wrubel J, Daubenmier JJ, Price CJ, Kerr CE, Silow T, et al. (2011). « Body Awareness: A phenomenological inquiry into the common ground of mind-body therapies. » Philosophy, Ethics, and Humanities in Medicine: PEHM. 6:6. doi:10.1186/1747-5341-6-6.
- 178.^Tuladhar R, Bohara G, Grigolini P, West BJ (2018). « Meditation-Induced Coherence and Crucial Events. » Frontiers in Physiology. 9:626. doi:10.3389/fphys.2018.00626.
- 179.^Goshvarpour A, Goshvarpour A (2013). « Comparison of higher order spectra in heart rate signals during two techniques of meditation: Chi and Kundalini meditation. » Cognitive Neurodynamics. 7(1):39–46. doi:10.1007/s11571-012-9215-z.
- 180.^McCraty R (2022). « Following the Rhythm of the Heart: HeartMath Institute’s Path to HRV Biofeedback. » Applied Psychophysiology and Biofeedback. 47(4):305–316. doi:10.1007/s10484-022-09554-2.
- 181.^Kaur J, Fahmy LM, Davoodi-Bojd E, Zhang L, Ding G, Hu J, et al. (2021). « Waste Clearance in the Brain. » Frontiers in Neuroanatomy. 15. https://www.frontiersin.org/articles/10.3389/fnana.2021.665803.
